|
THE PHYSICS
AND PATHOLOGY OF WOUNDS PART 3: CHRONICITY AND THE PHYSICS OF WOUND FAILURE Marc E. Gottlieb, MD, FACS Phoenix, AZ Revision 01-c, June
2, 2010, (Revision 01-a, February 10, 2010), Copyright © 2010 |
|
|
Preamble In Part 1 of this series, The Wound as a System and a
Controlled Machine, the main point was that the wound is not only a complex
system, but it is a non-linear controlled system. Control is the basis for all wound
dynamics, allowing the healthy wound to heal by one-shot dynamics, but
leading to complex patterns when wound healing is pathological. In Part 2, Auto-Immunopathy and the
Intrinsic Disease of Wound Healing, we went from a physics-engineering
perspective to a clinical-pathological one.
The general stroma, the auto-immune connective tissue disorders, and
the chronic wound were all equated through the principle of sustained chronic
inflammation leading to immune sensitization against stromal elements. Now, in Part 3, Chronicity and the
Intrinsic Disease of Wound Healing, we will bring together the engineering
aspects of the wound as a controlled process and the clinico-pathological
aspects of intrinsic auto-immune wound chronicity to understand why chronic
wounds fail to heal. This is a physics
based understanding that uses the principles of non-linear dynamics (the
physics of systems) to explain why the wound control loop cannot succeed in
the face of stromal auto-immunization and intrinsic wound chronicity. |
|
|
|
|
|
In Part 1 of this series, we looked at the wound as a system
subject to feedback and control. This
model of the wound can accommodate all conditions of wound physiology and
pathology, of normality, failure, and therapy. In Part 2, intrinsic wound pathology and
failure were examined, the condition of intrinsic degradation of wound
healing not attributable to extrinsic factors. This intrinsic disease of wound healing
results from the appearance of an abnormal population of chronic inflammatory
and immune cells which has complex disruptive effects on the two cell sets
which belong there, acute inflammation and wound module. These states have a critical association
with autoimmunity, microthrombosis, and other events which sustain
inflammation. From a biological perspective,
it is easy enough to see how the chronic inflammation might disrupt
healing. However, isn’t the purpose of
the Wound Control Loop to respond to and correct perturbation? There are reasons why the chronic wound cannot
respond as expected, and this is where the engineering science of Part 1 and
the biosciences of Part 2 meet to explain the physics of wound failure and
wound chronicity. |
|
|
|
|
|
The
wound is not just a tissue. It has
multiple substructures, a complex organization, and complex tasks. It is not a constituent tissue of some
other organ. It is itself an
organ. The wound is an organ. When we say “wound” here, we are not
talking about the injury nor the physical defect caused by the injury, but
rather about the wound repair process, the “wound module”, which is the
body’s response to the injury and defect.
It is a reserve anatomy and physiology, a transient ad hoc organ that
is triggered into existence by injury, then performs its appointed functions,
then wanes and disappears as its job is completed. Nonetheless, it has the structure and
functions that conceptualize any other fixed organ. So, we will start by comparing wound
healing to other organs. For any
organ, ask 3 questions: What
are its quintessential structures and functions? What
is the quintessence of its dysfunction or failure? What
is the fundamental physics relevant to its function? The
heart is a complex organ, with great complexity to its structure and
function. However, its quintessence is
that it is a pump with valves and pipes.
When it fails, it is an inadequate pump, damming the circulation. Some knowledge of fluid mechanics is
required to understand its basic physiology and pathology. The quintessence of the kidney is that it
is a filter and resorption membrane.
When it fails, it is tantamount to an occluded filter in any fluid
system. Basic hydraulics and ionic
chemistry are the core physical sciences that apply. The lung is essentially just a bellows and
diffusion membrane, and when faulty, ventilation and respiration are
impaired. Gas mechanics and diffusion
are the applicable physics. The eye is
a light collector and detector, its failure is blindness, and optics
apply. The ear is a sound transducer
and decoder, its failure is deafness, and acoustics apply. Skeletal structures are structural members,
their failure impairs support and mobility, and solid mechanics apply. Nerves are electrical control circuits,
their failure is equivalent to open circuits and faulty electrical
transmission, and electricity is the relevant physics. |
|
|
|
|
|
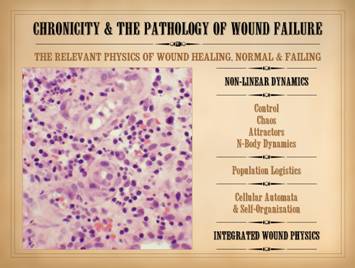
What
then is the quintessential structure and function of the wound? It is just a bunch of cells that get
together, make some stuff, then quiet down or disappear. What stuff are they making? They are making stroma. They are making, repairing, or restoring
the basic structural medium of the body, a composite material made of ground
substance, connective proteins, and blood vessels. They are making or restoring the basic
framing and utilities that the body needs to support all of the other
parenchymal cells and structures that have a more specific or parochially
defined role. Correctly stated, the
quintessence of wound anatomy (i.e. wound repair / wound module) is a
collection of mutually interactive self-organizing cell populations. What
is the quintessential derangement of intrinsic wound pathology and
chronicity? It is what happens when
this collection of self-organizing cells fails to organize to make new
stroma. It is what happens when they
fail to make new stroma due to their own disorganization and failed inter-operations,
rather than due to any extrinsically applied perturbation. New stroma depends on these cells getting
together into the right pattern and structure. If they fail to organize then the new
stroma is incorrect and cannot support associated parenchyma (like the
epithelium, and thus the wound does not close). In the words of its relevant physics, the
quintessential derangement or failure of this self-organizing system is that
it is a dynamical disorder of logistical self-re-organization among these
populations. What
are the fundamental or constitutive physical sciences – the physics –
relevant to the wound? The concepts of
self-organization, population dynamics, and mutual interaction are part of
the subject of non-linear dynamics, including control, chaos, population
logistics, and cellular automata. For
other organs, the relevant physics are mechanics, fluids, gases, optics,
acoustics, electricity, etc. For the wound,
it is non-linear dynamics. |
|
|
|
|
|
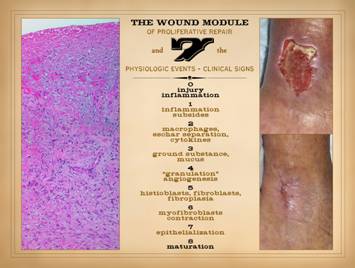
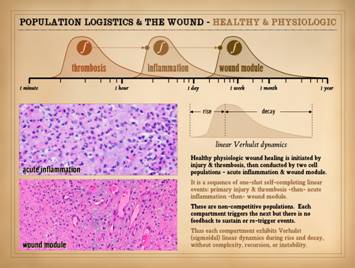
This slide comes from Part 1 of this series, “The Wound as a
System and a Controlled Machine”, slide 7.
It is meant to refresh a few points about basic wound anatomy and
physiology. On this slide, on the
right, a normal healthy wound goes from open to closed (contracted and
epithelialized) via the natural process of wound healing. Histologically, all of the reparative
events taking place in the wound have a well organized and recognizable
anatomy, and each of the features seen microscopically correlates with
something that is happening or can be observed grossly. Major events and correlated physical
findings are: 0 – Injury and
inflammation: They trigger the whole
process of response and repair. The
repair process is a reserve organ that arises only as needed. 1 –
Inflammation subsides: Acute
inflammation is the response to injury that includes initiating repair. However, sustained acute inflammation is
suppressive to wound repair. For
incidental injury, inflammation is a one-shot response that wanes as repair rises. As will be presented here, sustained
primary injury and sustained acute inflammation are one of the essential
dynamics required for wound chronicity. 2 –
Macrophages, eschar separation, and cytokines: Macrophages are transformed leukocytes,
arrived by inflammation, that have two major roles. Their afferent task is as phagocytic cells
to remove debris. This becomes
relevant to chronic wound pathology since this activity is a key step in
stromal auto-immunization. Their
efferent task is to initiate the repair process by the issue of proliferative
growth factors. 3 – Ground
substance and mucus: In advance of a formal
fibrous stroma or structure in the wound, aminoglycan ground substance is the
medium which early angioid cells need in order to migrate and function. 4 – “Granulation
tissue” and angiogenesis: Proliferative
angiogenesis creates the vascular network required for subsequent cells and
activities to function. Once this
occurs, fibroplasia and restoration of structural stroma can proceed. 5 – Fibroblasts,
and fibroplasia: Once angiocytes have
formed vessels within the aminoglycan layer, the environment now permits
fibroblasts to function. Fibroblasts
make the connective proteins required for a mechanically durable stroma. 6 –
Myofibroblasts and contraction:
To get the wound closed, specialized fibroblasts with muscle proteins
contract the wound, minimizing its surface area, lightening the load on
epithelialization. 7 –
Epithelialization: Final closure of the
wound is when epithelium is fully resurfaced, sequestering the mesenchymal
stroma underneath from the ambient environment. 8 – Maturation: Maturation is a long process in which the
excessive stroma of the newly healed wound remodels back to vascular and
connective protein densities and architecture which match normal dermis and
fascias. The vertical anatomy of the wound reflects timewise events and
sequences. The surface is happening
now. The fibroplasia layer deeper down
started so many days ago. Separation
of cells and populations (acute inflammation and wound module) by time and
strata are a crucial part of this physiology.
They become intermixed in the pathological wound. |
|
|
|
|
|
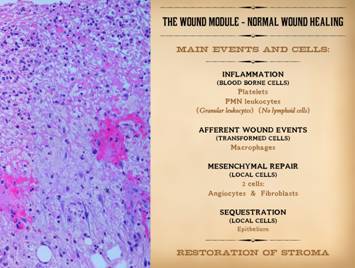
This slide also comes from Part 1 of this series. It is meant to remind that normal wound
healing has some general phases. The
response to acute injury is thrombosis and acute inflammation, dependent on
blood borne leukocytes. This
transitions into afferent wound events, mediated by transformed leukocytes,
which includes triggering the efferent reparative wound events. Repair is mediated by mesenchymal cells
with two – and only two – phenotypes, the angiocyte and the fibroblast. Wound closure, sequestration from the
ambient world by epithelial growth over the open mesenchyme, is the final
event. Several cells are crucial in
this whole process of repair: platelets, mononuclear leukocytes, macrophages
(transformed monocytes), angiocytes, fibroblasts, and epithelium. Acute inflammation, i.e. blood borne
monocyte-macrophages are the afferent population of wound cells. Wound module is the efferent repair
population made of local mesenchyme, i.e. angiocytes and fibroblasts. That is the crucial thing to remember here,
that wound healing events occur via two general populations of cells – acute
inflammation and wound module. |
|
|
|
|
|
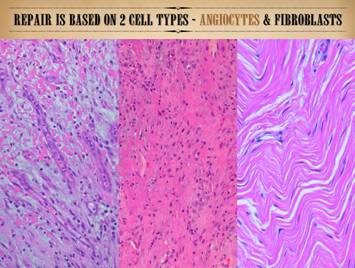
This slide is also form Part 1 of this series. It is meant to remind that the reparative
population, the wound module, is made of two – and only two – mesenchymal
cells, the angiocyte and the fibroblast.
They constitute the repair population.
Epithelium can also be considered part of the repair population, but
the mesenchymal component is what figures most prominently in the process of
stromal auto-immunization and the chaotic logistics of intermixed cell
populations 9as will be discussed in this Part). |
|
|
|
|
|
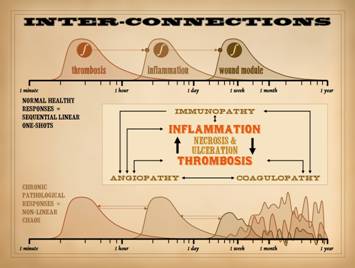
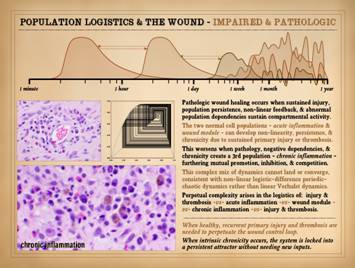
In Part 1, slide 32, and in Part 2, slide 46, we introduced the
concepts of sequential one-shot dynamics and their role in the normal
response to injury and wound healing.
We also hinted at the nature of chaotic dynamics, and how that arises from
abnormal feedbacks, dependencies, retriggers, or other sustentation of any of
the system sub-phases. We also looked
at how the co-dependent interplay between thrombosis and inflammation can
sustain abnormal dynamics and pathology, and how they are the input hook into
the system for primary pathologies to exert themselves on the wound, all with
necrosis and ulceration caught at the center of this. To refresh, recall the quintessential functions of inflammation
and thrombosis. How is injury
recognized? How is it cleaned up? How is the repair process started? Platelets-thrombosis are one pathway of
injury recognition. Once triggered,
they then initiate inflammation so the body can handle defenses, do damage
control, and then clean up. Thus, (1)
thrombosis triggers inflammation.
However, inflammation also creates a milieu that promotes thrombosis
via prothrombotic chemicals, leukocyte and platelet trapping, changes in
vessels and blood viscosity, etc. Thus,
(2) inflammation triggers thrombosis.
They trigger each other. This
complex non-linear system is self-amplifying.
In the case of single incidental injury, such as trauma, this
thrombosis-inflammation coupling ensures a swift ramp up of defensive
changes, but then the process subsides and settles, paving the way for
repair. In simple terms, trauma
induced thrombosis-inflammation is a one-shot. However, when there is repetitive or
sustained injury, then new thrombosis and inflammation keep getting
triggered, keeping the process alive or renewed. Chronic or repetitive injury,
thrombosis-inflammation triggered by a chronic hypercoagulable or
micro-occlusive disorder, and acute inflammation chronically induced by
immunity, allergy, or infection are the types of activities that perpetuate
these events – to the detriment of the host. It is the sustained interplay and mutual promotion of these
factors which disrupts the normal dynamics of healthy repair. Inflammation triggers thrombosis. Thrombosis triggers inflammation. Many other chronic inflammatory, immune,
and thrombotic disorders also trigger thrombosis and inflammation. Necrosis and ulceration are caught in the
middle, making more “wound”. The
orderly sequence of one-shot events is totally disrupted, becoming a
cacophony of feedbacks and retriggers that keep the
thrombosis-inflammation-repair dynamics in wild unpredictable states. As we will see here though, this situation
might seem wild, but that is how complex natural systems really behave. This is at the heart of why chronic wounds
fail, and these dynamics are understandable by studying their relevant
physics, non-linear dynamics. |
|
|
|
|
|
As we start to explore the physics and pathology of wounds, keep
the following quintessential points in mind: The wound module is a special ad hoc reserve organ. It is a collection of mutually interactive self-organizing cell
populations. The wound is not a pump
and pipes like the heart, not a diffusion membrane like the lung, not a
structural member like a bone, not a light collector like the eye, and not
like any other already organized organ with an ongoing specific function. It is a collection of mutually interactive
self-organizing cell populations. It
has no other function than to get organized (into a generic stroma that is
the foundation for other tissues and organs.) Wound failure is a dynamical disorder of logistical
self-re-organization among the wound’s constituent cell populations. When it fails, it is not an inadequate pump
like the heart, not an inadequate filter like the kidney, not an inadequate
bellows or diffusion membrane like the lung, and not like any other organ that has ongoing specific function. When it fails, it simply fails to get
organized into its intended final form, to complete its task to become
something and then cease. The relevant natural science which explains its quintessential
structure, function, and dysfunction is physics, specifically, non-linear
dynamics, including control, chaos, population logistics, and cellular
automata. The relevant physics is not
fluid dynamics as it is for the heart, not optics as it is for the eye, not
acoustics as it is for the ear. It is
the science of populations and the principles of their interaction, control,
and self-organization. When it fails, it is, in the terms of its relevant physics, a
dynamical disorder of self-organizing populations. |
|
|
|
|
|
The wound, the organ of repair, is anatomically and functionally
a collection of cells trying to organize themselves. The relevant physics is Non-Linear Dynamics
(NLD), the study of complex systems, which applies to the normal healthy
wound and the failing wound. The broad
subject of NLD has a variety of components.
Those that will be studied here are in 3 areas: (1) the general timewise behaviors of
complex systems, including control, chaos, attractors, and N-body
dynamics; (2) population logistics,
the study of collective and competitive group behaviors; (3) cellular automata and
self-organization, the study of how deterministic rule-based systems assemble
themselves with minimum information and maximum efficiency. The purpose of all of this is to explain
why impaired, failing, non-healing wounds are that way, for reasons that go
beyond the conventional bio-pathology of the system. |
|
|
|
|
|
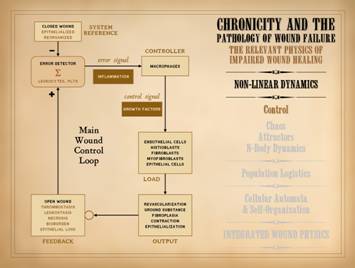
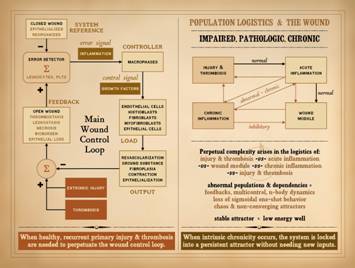
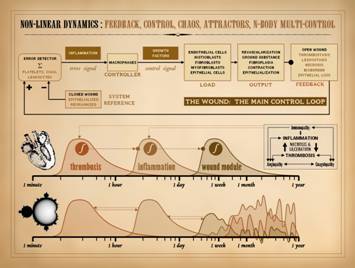
The subject of control was covered in Part 1. of this series, “The
Wound as a System and a Controlled Machine”.
The main wound control loop is shown here, and needs no further
comment. However, it is important to
understand the connection of control to non-linear dynamics. Control means feedback, to report the state
of the system so that errors can be corrected. Non-linearity is explicitly defined by
feedback, the system state or output at one moment becoming the input into
the system in the next moment.
Controlled systems are ipso facto non-linear systems. The wound is a non-linear system. The control within that system ensures
that, when healthy, injured tissues return to a state of restored stromal
architecture and integrity. |
|
|
|
|
|
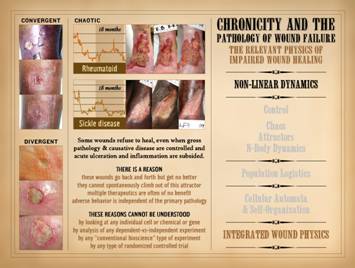
Note on slide 7 that feedbacks and sustentation of primary
system triggers could lead to seemingly erratic behavior, aka “chaos”, which
for the wound implies non-healing. The
subjects of chaos, attractors, and N-body dynamics explain such complex or
unpredictable timewise behaviors in non-linear systems. Non-linear dynamics is a modern subject,
relevant only since the 1970’s – 1980’s, after the advent of digital
computing. However, the basic concepts
of the subject and the inherent need to have the subject have an important
history over the past 300 years. For the past 3 centuries, our main system of technical and
engineering mathematics has been the calculus of Sir Isaac Newton (1643 –
1727), the mathematics of motion and change.
The calculus lets you create analytical (linear) equations that
describe the state of a variable versus another variable. The importance of Newton’s mathematics
cannot be overstated, but the calculus does not solve all problems. Newton himself knew this perfectly well. His gravitational equation allows the
behavior of two mutually interacting bodies to be described with exact
precision, e.g. position, energy, velocity, momentum versus time. The problem is that the simple analytical
equations which describe two mutual bodies do not exist for describing 3 or
more mutually interacting bodies. Imagine
for instance that you need to compute the orbit of a large moon m
around planet p. Easy enough
with calculus, and the results can be expressed directly as m = f(p),
or parametrically as m = f(t), p = f(t). But what if it is a trinary system, with
another large moon n. In that
case, m = f(p), n = f(p), and n = f(m). These three mutually interacting bodies
cannot be solved with Newtonian calculus, i.e. there is no general solution
in the form of a functional equation.
This dilemma is known as the “3-body problem”, generalized as the “N-body
problem” for any number of mutually interacting objects. For 3 centuries, scientists and mathematicians were instructed
to stay away from such problems as “intractable”, and engineers had to depend
on approximations and limited grainy iterations to solve real world
computations. However, by the end of
the 19th century, mathematicians were starting to come to grips
with the fact that complex real world systems might not be solvable with
linear analytical equations, but that the real world is the real world, and
that mathematics would have to rise to the challenge and find methods to
describe reality. Some of the most
important insights into this issue were made by French mathematician and physicist
Henri Poincaré (1854 – 1912) who made seminal contributions to the subject of
3-body problems, and in so doing became the first to “discover” or anticipate
the concept of deterministic chaos.
The problem then though was that the iterative solutions to such
problems could hardly be visualized, because such solutions depend on
numerous repetitive calculations. His
work, and that of other notables such as Gaston Julia (1893 – 1978) lived in
abstraction and theory until automated computing in the latter 20th
century permitted the direct calculation and visualization, the “solution” of
such problems. In the biosciences, we continue to live with the legacy of
“intractability”. For a century and a
half, experimental physiology and biosciences have been mired in the concept
of linear models of dependent-versus-independent parameters in an otherwise
invariant environment. This approach
to research has characterized the linear relationships of millions of
cellular and chemical interactions – all crucially important, but none of
that describes how systems as a whole behave (more on this on slides 15 &
16). In this, the “century of the
system” (see Part 1, slide 5), and with the tools to solve iterative N-body
systems, complex biosystems need to be approached for what they are – complex
non-linear systems. Physicists,
chemists, earth and atmospheric scientists, meteorologists, geologists and
geographers, and scientists from most of the physical and earth sciences have
been doing this for 20 – 30 years now.
But the biosciences are largely stuck in 19th century
methods of investigation – ironic in that biosystems are far more complex and
in need of such analytical understanding more than most other physical
systems. The dynamics or behavior of
wound healing, especially the impaired or failing wound, is a perfect example
of a complex non-linear system.
Classic bioscience experiments cannot explain why a wound fails,
because these are problems of complex systems, for which physics – non-linear
dynamics – is necessary to understand the real failures of the system. As is presented here, the failed wound, a
complex non-linear system, can be understood by a several aspects of
non-linear dynamics, not the least of which is that the system fails under
the confluent interaction of three “bodies”, in which each body is its own
complex element, a non-linear “population” of cells. It is all “intractable” according to
“classic” physiologists, but it is the real world with real explanations
based on contemporary physics. |
|
|
|
|
|
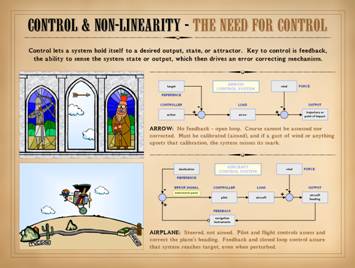
This slide is also form Part 1, Control . . . It is meant to remind that biological
systems have control, allowing them to “steer” toward a desired state or
system reference. An open loop system,
like the arrow, requires calibration, and calibration can be upset, making
open loop systems prone to error at the output. Closed loop systems, like the airplane,
which can sense and correct errors, are more fault tolerant, and they can
meet their mark even when perturbed. Consider for a moment that some space alien is here on a field
trip to study us. He observes without
interacting, privy to no knowledge or communication other than what he can
observe visually from his vantage point high in the sky. If he were to observe the airplane and
pilot, what would he see? He might see
a straight line of flight, from origin to destination. “Might”, because a perfectly straight path
would be contingent on no winds, perfect navigation and flight control,
etc. A pure straight line is unlikely,
but nonetheless, the flight path will likely be nearly straight – a few minor
twists, a curve here, a variance there, but still more straight than not,
with a small error if fit to a regression curve. The space dude could figure out easily
enough that this is a controlled flight, that variances are due to
perturbations like wind or whatever, and that the pilot or system is
controlling off of some basic concept of efficiency based on optimizing
economy or energy on the way to the target.
The deterministic rules behind the control of this system would be
easy to infer, especially since the space dude himself is technologically
savvy. In fact, if the space observer
had sensors to observe wind velocity
and direction, he could correlate those parameters with the subsequent responses
of the airplane. Soon enough he would
have all the data necessary to understand the deterministic rules of
response, then model the system well enough to accurately anticipate the
responses of the airplane to changes in the wind. |
|
|
|
|
|
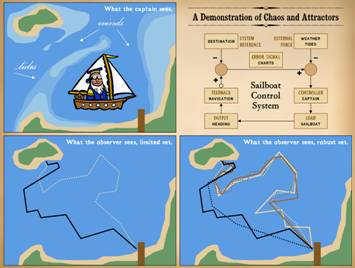
Control, determinism, and analytical predictability were all
evident in the airplane control system.
Now, we will look at a sailboat, in which control and determinism are
equally strong, but with a more complex set of rules and responses, enough
that the system state or output is not analytically predictable – aka
“chaos”. The sea captain is an experienced old salt who ferries back and
forth between the mainland and the island. He has his nautical charts and an implicit
knowledge of the underwater terrain.
He is aware of hazardous shoals, reefs, and sandbars. He also knows of a shallow channel shortcut
that is passable at high tide but must be avoided at low tide. He understands the wind and his sails, and
he has complete mastery of the craft.
Every trip he makes is perfect on points of safety and economic
efficiency. On the lower left, the captain has made one round trip, sailing
out at high tide through the shallow channel, then taking the long way home
at low tide. Tack lines vary, based on
wind, water, and hazards. What would
our friend the space alien see if he was observing? He might not figure this one out. He might come away downright confused about
how the boat operates – i.e. what are the rules of control. First, consider that the space dude is NOT
technically savvy about boats. Perhaps
their planet has no large bodies of water, so those technologies never
developed. Or maybe, he is an inland
city boy on his home world and never took a ride in a sailboat himself. If so, he would not understand how and why
the boat must tack if it is to sail into the wind. Perhaps his observational platform up there
cannot sense underwater forms (sandbars) nor sea height (tides) nor the wind. Thus the space alien is unaware of these
rules and perturbations. All he can
see is the mainland, the island, a featureless surface between, and the boat
shuttling back and forth. If the space
dude assumes that the boat should move in a straight line for the sake of
economic efficiency, then the strange patterns he observes will make no
sense, especially if he observes just once.
Yet the system is not without rules, so it would seem, because this
seeming nonsensical and highly erratic not-a-straight-line-shortest-distance-between-two-points
pathway nonetheless eventually converges back at the starting point. If he took the time to observe many transits, and had a robust
data set, could he then figure out the rules?
No. He could theorize or
hypothesize, but unless he gets direct knowledge of the underwater geography,
winds, tides, and how a sailboat operates, he cannot figure out the
rules. This is especially true since
the response to a nearly identical set of conditions may vary, for instance a
few degrees change in the wind or a week’s change in the lunar phase may make
the captain take a completely different course. From the captain’s point of view, from the
operational physics point of view, this system is strictly
deterministic. The captain will make
the same exact choices, time after time, based on explicit rules of safety
and efficiency, for any given set of inputs.
It’s just that the inputs are many and variable, and small changes may
make a big difference in subsequent responses or patterns. If the space alien is unaware of the small
perturbation which triggered a big variance in output, then the rules or
responses will seem arbitrary. If the
space alien makes a robust set of observations, what he will learn is that
the boat sails on an “attractor”, a state space of permissible values or
allowances. Where the boat will be at
any given time is not strictly knowable absent knowing all of the rules (this
is the principle known as Laplace’s Demon).
The captain knows all of the rules.
The space observer knows few of them, so he will have to infer what he
can from the shape of the attractor.
The complex or seeming erratic pattern of the attractor becomes very
simple once you know the rules, but alas for our space friend, all he
observes is apparent mayhem. This is
the concept behind mathematical chaos.
“Chaos” in the vernacular sense implies mayhem and arbitrariness, but
in physics-nature-math, chaos is the antithesis of mayhem. It is just the complex behavior of complex
or non-linear systems driven by strict deterministic rules, whether we know
or not what those precise details or rules might be for a given system. |
|
|
|
|
|
. . . All of which brings us to the point of understanding what
non-linearity chaos, and attractors really are. Keep in mind that this is how all real
world complex systems operate. The mathematics
on these few slides is not meant to intimidate or overwhelm – in fact, just
the opposite. These abstract
mathematical larks are a simple way to illustrate how complex real world
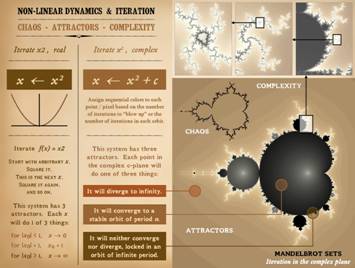
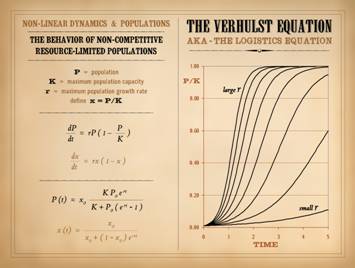
systems behave, starting with iterations on x2. The map (graph) of y = x2 is a simple
parabola. This is a linear function, a
proper equation of algebra and calculus, continuous, differentiable, with a
one-to-one (and “onto”) relationship between the input (independent variable)
and the output. Now, let’s iterate x2. xn+1 ! xn2. Pick an arbitrary value for x. Square it,
Make the answer the new value of x, then square it, and so on. What will happen. If the start value is x = 1, then the
function will converge (immediately) on the value of 1. If first x is less than 1, then repetitive
iteration will drive x towards zero, converging at 0 after infinite
iterations. Likewise, if first x is
greater than 1, then x converges on infinity.
X has 3 attractors: it
converges to 0, lands on 1, or diverges to infinity. The fact that the system depends on itself,
that its own value or output feeds back as the input on each iteration, this
is what defines it as a non-linear system.
Simple enough. However, even
the seemingly mundane as y = x2 can become very complex
when you throw in a few extra rules. The “extra rule” here is that we will iterate x2 in
the complex plane. Recall that complex
numbers have a real part and an imaginary part (i = square root of -1), and
they must be represented on a rectilinear graph. So as to avoid the trivial attractors of
zero-one-infinity, we will add a constant c, so that we are now iterating xn+1 ! (xn2 + c). We will map the iteration in the complex c
plane, meaning that every point on the grid (or display screen) is a value of
c. We can then pick an arbitrary
starting value of x, then iterate the equation. Depending on how the iteration behaves, we
will assign a certain color to each c point or display pixel. Specifically, we will observe the values of
x as we do the iteration, looking for the signs that it is going to diverge
or converge. We pick an arbitrary
discriminator, such as 10. As we
iterate, if x exceeds that value, that is taken as a sure sign that the
function will diverge, on its way to infinity. We then assign a color to that display
point based on how many iterations it took to exceed the divergence
discriminator. We are also looking for
repetitions in the values of x as we iterate it. If a value repeats itself, that means a
closed orbit has been achieved (and it will continue to repeat itself). We count how many iterations are in the
orbit, then assign a color value based on that number. When we are finished iterating each point
in the display field, for each value of c, what do we get? We get the amazing structure shown, the
Mandelbrot set of iteration in the complex plane. Points within the Mandelbrot set are convergent – they have
settled into a repeating orbit. The
closer they get toward the center, the smaller the orbit, the fewer
iterations it took to find stability.
Points outside are divergent – they are “blowing up” to infinity. The farther away, the faster the values are
diverging, and thus the fewer iterations needed to cross the “escape”
discriminator. The color bands
visually clarify the structures and relationships. What is happening in between the convergent
and divergent zones? If we are outside
in the divergence zone, we find that the farther out we are, the fewer
iterations needed to diverge. The
closer in we are, the more and more iterations required to diverge,
approaching an infinite number of iterations at some limit. If we start off inside, in the convergence
zone, we find that if we are close to the center, then orbits converge after
just a few iterations. As we move
outward, it takes more and more iterations to converge an orbit, approaching
an infinite number of iterations at some limit. The two zones, divergence and convergence,
both take an increasing number of iterations to not do their thing as they
move towards each other. At some point
it takes an infinite number of iterations to find an orbit (no repetition of
values). At some point it takes an
infinite number of iterations to not diverge (remaining in bounds at low
values). These limits of infinite
iteration are the boundary between the inside convergent zone and the outside
divergent zone. This transcendental
boundary of infinite orbit is the “chaotic” set of this function. This boundary is of infinite complexity,
which can be seen by zooming in, as shown on the upper panels. The point is that this seemingly mundane system, a simple
parabola or second order equation, can yield infinite complexity when the
problem becomes non-linear. This
function is highly deterministic – rule based – as all non-linear systems
are. Yet the value or state of the
system can be very irregular and unpredictable – aka chaotic – which leads to
complexity. Even so, values will tend
to be in certain permissible zones – aka attractors. The Mandelbrot set of iteration in the
complex plane is a very handy way to visualize these concepts, but these are
the inherent principles of all non-linear systems, including most biological
systems, including the wound. The
Mandelbrot set per se may not show up in your wound, but principles of
non-linearity, attractors, divergence, convergence, and chaos are there all
of the time. |
|
|
|
|
|
The principles of non-linearity, iteration, control, and chaos
must now be extended to the subject of N-body problems and
multi-control. The concept of the
N-body problem was introduced on slide 11.
It should be intuitive that this is relevant to the wound which has
“countless” numbers of interacting cells and chemicals. To understand 3-body problems, consider the
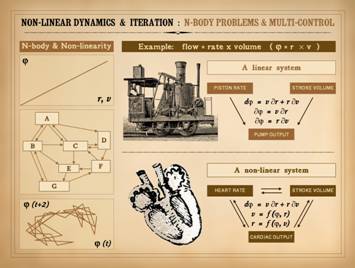
functions of a pump. The antique
locomotive runs off of a steam pump.
The pump has a certain flow rate, e.g. gallons per minute. The flow
(p) is a consequence of the stroke volume (v) and the stroke rate (r),
specifically p = vr. Engineered
machines of this sort can be made so that stroke volume and stroke rate can
be adjusted by the operator independently of each other. Each is an independent variable of the
system, neither contingent on the other.
Thus, p = f(v) and p = f(r), but v & r
have no dependencies. The total flow
or output varies as v and r vary. If v
and r vary concurrently, you can still directly calculate variations in flow
by prorating the contributions of each independent change. This is the purpose of partial differential
equations: dp = v¶r + r¶v. This is a simple analytical non-linear system. Its output can be functionally
graphed. Simple. Non-linear. Next, Consider the heart.
It too is a pump. Flow (aka
cardiac output) equals stroke volume times heart rate, p = vr, just
like the steam pump. But, there is a
crucial difference between the two pumps.
In the heart, as in the steam pump,
p = f(v) and p = f(r), but also v = f(r). Consider a basic point of physiology: as
heart rate increases, stroke volume can diminish because diastolic filling
time is impaired, i.e. volume and rate are contingent. Another point: as stroke volume increases
(as during exercise, due to increased venous return), ventricular stretching
drives a higher heart rate, i.e. rate and volume are contingent. Each is a function of the other. There are three “bodies”, three independent
variables, three mutual sets of functional dependencies. As a 3-body problem, there is no
constitutive general solution to this system that lets you functionally map
any one versus another nor versus time.
If you want to experiment with this system, you have to iterate it. In complex non-analytical systems, such as 3-body problems, you
simply cannot state the value of a dependent variable as a function of time
or some other independent variable.
All you can do is say that y = f(x), x = f(z),
and z = f(y). This
system of three linear equations can be “solved” only with iteration. What does that mean? The method to solve the whole system is to
recognize that over infinitesimally small intervals, y = f(x)
is valid unto itself, and z need not be accounted for – and likewise for the
other 2 equations. Thus, in each
go-around, you take the current values of each variable and use them to calculate
each of the three linear equations as though they were independent of each
other. Then you reiterate, taking the
new values of each variable and redoing the calculations. The thinner the timeslices, the more
calculations you do, and the more accurate your model is (the set of
equations and their one-versus-another linear characterizations), then the
more accurately the iterations will mimic-model-recreate real world dynamics. Each time you iterate, you are plugging the system values back
into the system operations. This is
the very definition of feedback, which ipso facto makes this a non-linear
system. Such systems generally have
ways to sense and monitor the feedback and compare the system state to a
reference. The train engineer knows
when the engine is running too fast or too slow and can make suitable
adjustments. The heart “knows” when
system performance is creeping out of permissible bounds and regulates things
back in bounds. As we saw in Part 1,
overwhelmed control or bad control can occur, but when system components are
behaving properly and not over-stressed, the machine as a whole should be
performing properly, “physiologically”.
The heart of course has plenty more parameters that must interact than
just rate-volume-flow, making it a “bazillion-body” system. The same is true for the wound. Yet within these complex systems, there are
subsystems and embedded loops which each have their own feedbacks,
dependencies, and regulated control.
For example, in the wound, the thrombosis-inflammation loop exhibits
control. The thrombosis, inflammation,
and repair one-shots all have control.
Even the way in which an angiocyte migrates then reassembles in
response to a gradient field of VEGF is a strictly reference-driven
controlled process. All of these
events have “hooks” into the other control loops, acting as extrinsic
promoters or inhibitors. This matrix
of interconnected control loops is “multi-control”. As we have already seen, when a well
behaved simple system should be acting like a one-shot, but various forces
and perturbations keep elevating or rocking the system, then chaotic dynamics
should ensue. With complex
multi-control, chaotic dynamics is largely assured. Remember to keep in perspective the difference between the
layman’s vernacular meaning of “chaos” versus the technical meaning. The description of multi-control leading to
chaos might sound awful to the naive observer, but in complex physiological
systems it is a cornerstone of healthy physiological function. Multi-control is what keeps physiological
systems in bounds. The
moment-to-moment variability of a given parameter might seem erratic,
unpredictable, and non-analytical, but the healthy system will keep that
parameter within permissible bounds, within a certain “state space” or
attractor. You might not know
moment-to-moment what the next value or variation of a variable will be, but
you will know that it stays on its attractor, unless it gets sick. Chaos, in the technical sense, is simply
the way that non-linear systems will behave.
There can be no other choice, no other way about it for N-body
multi-control systems. The more layers
of control, the more non-linearity, the more erratic the chaos, but that is
the way that healthy complex systems behave.
Chaos in physiological systems is not only normal, not only healthy, but
if it disappears that is bad. Continuing
with the heart analogy, the work of cardiologist Ary Goldberger is noteworthy
(e.g. Goldberger AL. Nonlinear dynamics, fractals and chaos: applications
to cardiac electrophysiology. Annals Biomed Eng, 1990, 18:195-198.) As a complex multi-control non-linear
engine, heart parameters should be chaotic.
If the beat-to-beat interval of the heart is finely measured, there is
plenty of erratic fine variation, and this can be shown by several
mathematical tools to be not just chaotic, but a consequence of control (such
as the embedding diagram, left bottom).
In sick patients with terminal heart disease, the beat-to-beat chaotic
variability disappears shortly before cardiac arrest and death, heart rate
instead becoming an exact fixed rate, meaning periodic or harmonic. Harmonic periodicity is a sign that many
layers of control have gone off line.
“Chaos is good, and well-behaved periodicity is bad” . . . the physics
of complex systems may seem a bit counter-intuitive if you have not studied
the subject. However, that is the way
that healthy complex systems are supposed to behave, all a consequence of
multi-control. The straight line in the left upper panel shows a typical linear
graph, such as (pump flow) = (proportionality constant) x (rate), the kind of
analytical function that has governed biological research for so long. The left middle panel shows the kind of
interconnection diagrams (see next slide) that characterize actual complex
biological systems. Several nested or
interlocked loops can be seen, and these multiple dependencies are the basis
for multi-control, which will lead to chaotic dynamics. The left bottom panel is typical of the
kind of mapping that must be done to see the structure in chaotic data sets,
in lieu of a functional plot such as in the upper panel. There are many ways to demonstrate chaotic
dynamics and attractors. The method
illustrated is an “embedding diagram”, in which one variable is plotted against
itself, its current value versus its value x number of iterations hence. This of course is most appropriate for
non-linear systems that feedback and depend on themselves. The embedding diagram assumes that with
feedback and control, the value of the system a short time from now will have
a functional cause-and-effect relationship to its value now, based on its
deterministic rules and physics. In
making such plots, structure and attractors can be revealed. The wound is a perfect example of complex multi-control. Chaotic dynamics are normal, but when the
system is healthy, feedbacks and dependencies are minimized, and each phase
or component of the process can run its one-shot event, the overall process
likewise being relatively smooth and regular as it returns to the system
reference “baseline” and a healed wound.
When pathology and abnormal dependencies occur, the system develops
abnormal degrees of feedback, multi-control, and n-body dynamics. This may be counterproductive or
detrimental from a clinical point of view if the wound becomes chaotically
locked into an orbit and doesn’t want to heal, but this is all normal and
expected behavior from a physics point of view. |
|
|
|
|
|
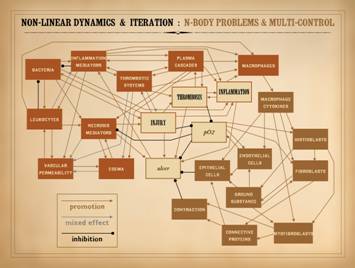
Perhaps you remember from medical school that giant chart of the
metabolic pathways. Undoubtedly you
have seen other highly interconnected block diagrams, such as the one shown
on this slide, in a variety of lectures or textbooks, concerning a variety of
topics in physiology or pathology.
These charts always seem to elicit a certain mood of tongue-in-cheek
irreverence, or a certain degree of apology or cynicism or whatever. There is a sense that although the author
is clever enough to chart the inter-connections, that no sensible or dutiful
post-doc would ever actually design an experiment based on such a jumble, nor
would any self-respecting bioscientist ever attempt to derive from such a
mess a unification theory that would explain integrated metabolism or
pathology based on such a briar patch of blocks and arrows. Yet this is the real world, real physiology
based on real dynamics. When a wound fails,
or there is unexpected worsening or improvement, or there are contrary
responses to therapy, the reasons need not be a matter of confusion. The understanding of such situations is
built into these interconnection diagrams, once you start to understand how
the elements in such systems inter-operate. In the example shown, a variety of elements relevant to wound
physiology and pathology have been inter-connected. This is non-rigorous, with an arbitrary
selection of relevant (but generally important) elements. Red items pertain more to injury, acute
inflammation, and afferent wound events.
Brown blocks pertain to efferent or reparative events. The ivory blocks in the center are major
states or common elements. They have a
multitude of interactions, either promotional-stimulatory versus inhibitory
versus mixed effects depending on circumstances. The numerous blocks qualify this as an
n-body system. The varying patterns of
feedback create complex multi-control, with individual loops nested, intersected,
and inter-connected. Classic
bioscience experiments have studied and published the parameters, equations,
and coefficients that characterize the interactions between any pair of
elements. Each 2-body pair has a
strictly deterministic profile. But as
was seen on the last slide, the mutual inter-dependence of many elements in
this system means that system state or output will have complex
non-analytical non-functional non-harmonic patterns, i.e. chaos. This graph was made arbitrarily – relevant but generally
important items were selected until space ran out. Then, the blocks were connected to others
based on common knowledge and principles of inflammation, wounds, cell
biology, etc. But in looking at the
overall result, notice something interesting.
There are few inhibitory lines.
In fact, the three early state events – injury, thrombosis,
inflammation – have no inhibition whatsoever.
This was not an intentional design goal, not a deliberate artifice,
not a rhetorical device to prove a point.
The control elements were interconnected according to basic knowledge,
and guess what – no suppression of injury, thrombosis, inflammation. We can suppress them therapeutically, but
the natural system needs no suppression.
These events extinguish themselves – they wind down and dissipate on
their own as long as they are not re-triggered or flared up. This dynamic sits at the heart of why
healthy wounds heal, and why they do not when wound chronicity sets in. |
|
|
|
|
|
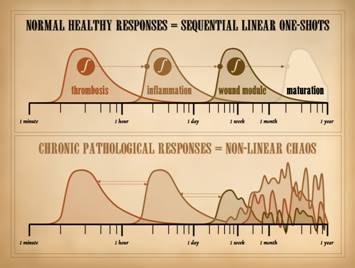
This slide comes from Part 1. of this series, “The Wound as a
System and a Controlled Machine”, slide 32.
In looking at wound repair as a controlled machine, we saw that when
the system is healthy, the tissues can return to baseline status, i.e. heal,
along a smooth trajectory that indicates good control. If we are more precise and look at the
distinctive states of the wound – injury (then) thrombosis
(then) inflammation (then) wound module (then) maturation – we see that each
state has a smooth ramp up then decay, a one-shot. Each state is well controlled. Each state decays and dissipates as its own
respective control loop succeeds in restoring its own sub-system to stability
or normality. Direct suppression or
inhibition from the outside is not needed.
This is inherent in the principles and mathematics of control. This is how the healthy wound behaves, from
injury to healed. We also introduced the concept that each of these major states
acts as some sort of trigger (generally an integrator or threshold switch) to
turn on the next major event in the
sequence. As long as the system is
healthy and nothing re-triggers it, then these major states act as a series
of one-shots, separated sequentially in time, and even separated in space, as
we saw in Part 2, “Auto-Immunopathy and the Intrinsic Disease of Wound
Healing”, slide 52. However, under
conditions of repetitive or sustained injury or pathology, then abnormal
feedbacks, retriggers, or other sustentation can occur, either sustained
forward-stimulation of the downstream event, or backward-stimulation of more
acute events. In Part 2, we saw the
various reasons why the pathological wound is subject to abnormal
sustentation and perpetuation. These
conditions of sustained injury or disease result in feedback, multi-feedback,
and n-element interactions. The
dynamics of the output will seem erratic and unpredictable, even though they
are highly deterministic interactions – i.e. chaos. We have now seen here in Part 3 what is
really meant by chaos and how it occurs.
It should be no surprise then that wound dynamics will be chaotic when
anything upsets the orderly series of one-shot events that characterize
straight-and-narrow healthy wound healing in healthy systems. |
|
|
|
|
|
The main wound control loop reduces the wound module to its
quintessential dynamical elements, making the principles of feedback and
control, the basis for non-linearity, easy to see. The Mandelbrot set of iteration in the
complex plane is an easy way to illustrate the complexity or chaotic behavior
that can arise from very simple rules and recursion (feedback). In complex physiological systems, the
elements of feedback and control, non-linearity and complexity become more
elaborate and multi-layered, such as for the heart where even a rudimentary
concept like pump output becomes a three-body problem. What it all means is that absent a detailed
knowledge of all rules and parameters, it is impossible to predict or
calculate with exact precision what the values of parameters will be in
moments to come. Yet it is easy enough
to know where the general attractors of the system are, i.e. what its
permissible state space is, and what the general dynamical behaviors or
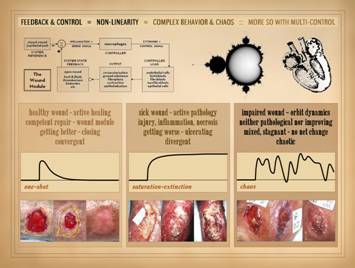
responses of the system will be. For the wound, with all of its complex physiology, yet with
basic feedback and control, its state space can be reduced to three
attractors: the healthy wound, the
sick wound, and the impaired wound.
For each, there is: (1) a tangible or gross state that can be
observed; (2) a physical state of the machine and its parts; (3) a teleological
state, a state of intent or goal or heading; (4) a dynamical state, the
activities or state of the control loop.
From this point of view, the three attractors can be characterized as: The healthy wound.
Tangible state: the wound that
is actively healing. Physical
state: the wound module is healthy and
competent, functioning properly. Teleological
state: the wound is getting better and
closing. Dynamical state: the wound is convergent, it is settling
toward a stable or resolved state. The sick wound.
Tangible state: the wound is actively
pathological, feeling the effects of active injury, disease, thrombosis, and
induced or applied inflammation.
Physical state: active injury
and ulceration are occurring, with inflammation-lytic or thrombo-infarctive
forms of necrosis. Teleological
state: the wound is getting worse,
actively ulcerating and getting bigger.
Dynamical state: the wound is divergent,
“blowing up” to large values of area and pathological histology. The impaired wound.
Tangible state: the wound is in
a go-nowhere state of incidental back-and-forth with no real progress,
“orbiting” in the same sense as the underlying dynamics. Physical state: the wound is neither pathological nor
improving, i.e. disease (the sick divergent wound) is arrested but the wound remains
not healing. Teleological state: the wound is neither getting better nor
getting worse, with mixed features from one observation to the next, but
overall stagnant with no net change over long intervals. Dynamical state: the wound is chaotic, on the cusp between
divergence and convergence, orbiting endlessly until some strong perturbation
can knock it to one side or the other. |
|
|
|
|
|
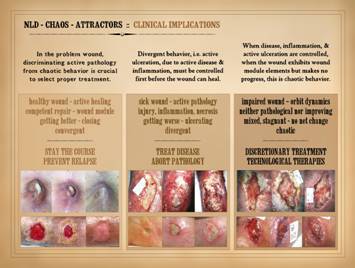
On the last slide, we summarized the implications for the wound
of non-linear dynamics, chaos, and attractors. Here, we can translate that into clinically
relevant information relating to wound diagnosis and therapy. Remember, the wound can be in one of three states. It can be healthy and getting better. It can be sick and getting worse. It can be impaired and just not going
anywhere, no longer sick but not yet getting better. Proper diagnosis is the keystone to all good care. Without a correct diagnosis, proper
treatment cannot be selected nor managed.
Among the various constituent elements of a thorough wound diagnosis,
one is the dynamical state of the wound, and the clinician MUST be able to
distinguish or discriminate active pathology from chaotic behavior. We have at our command many therapeutic
tools to treat problem wounds, Some
are tools to treat underlying disease, and some are wound therapies to
stimulate an impaired or retarded wound.
Without discriminating active disease from dynamical “lethargy”, wrong
treatments will be selected and care will be wasted or the patient
harmed. For example, in the actively
pathological sick wound, applying expensive stimulatory therapies such as
recombinant growth factors or cultured living cell products in the face of
active disease and acute inflammation is a profound waste of time, money, and
resources. For the chaotic impaired
wound, boosting steroid doses (e.g. for a rheumatoid wound) after
immunopathic inflammation has already subsided is wasteful and potentially
harmful. Divergent behavior, i.e. active ulceration, due to active
disease & inflammation, must be controlled first before the wound can
heal. When disease, inflammation,
& active ulceration are finally controlled, and the wound starts to look
healthy, but it then makes no progress, this is chaotic behavior. These two states, divergence versus chaos,
active disease versus non-healing, must be discriminated in order to choose
correct treatment. Most experienced
wound clinicians understand this implicitly.
It is the purpose of these papers to clarify that when the wound
should be healing but isn’t, that this is a thermodynamically stable state,
and there is a basis in physics for this situation. Understanding this is necessary to have
greater equanimity in your approach to the wound and the patient, and to
become more exact in choosing treatment. Remember, as an organ the wound is just a collection of cells
doing interactive things. If the
correct treatments are to be chosen, then its diagnoses must reflect the
behavior of cell populations – i.e. non-linear dynamics and population
logistics. It is the same as the
diagnosis of heart failure (pump with valves and pipes), where valvular
dysfunction, ventricular diameter, wall motion, ejection fraction, diastolic
dysfunction, cardiomyopathy, rhythm, and cardiac “fuel” (coronary supply) are
all crucial to the thorough diagnosis and selection of treatments. For the cardiologist, nuanced differences
in these parameters can make differences in choice of treatment. For the wound physician, understanding the
dynamical states of cell populations is required to understand the status of
the wound and pick proper therapy. For
the three general attractors or states of the wound, the following are the
major therapeutic imperatives: The healthy wound.
(Active healing, competent wound module and repair, getting better and
closing, convergent.) “Stay the
course.” Continue current therapies,
and make sure that there is no relapse or recurrence of primary disease or
injury. Therapies may be basic and
passive, non-specific general care to keep an intrinsically healthy wound
healthy, or they may be discretionary or technological therapies that have
successfully turned the wound from an impaired non-healing state to its
current healing status. The sick wound.
(Active pathology, with injury-inflammation-necrosis, getting worse and
ulcerating, divergent.) Treat the
active disease or injury, and abort the pathology. Ameliorate or mitigate additional risk
factors and stressors. Get the wound
under control, without active inflammation, necrosis or ulceration, without
symptoms nor threat to general health.
Deliver the wound to a state of healing (the healthy wound) or at
least neutrality (the impaired wound). The impaired wound.
(Orbit dynamics, neither pathological nor improving, mixed-stagnant
with no net change, chaotic.) This is
the time for discretionary treatment, the time to select specific and
directed therapies meant to force the wound to close or to implement some
other definitive plan of care. At one
end of the spectrum of options, this might be a deliberate choice to accept
the impaired status of the wound and treat it chronically as open, with basic
hygienic care to avoid relapse. At the
other end of the spectrum, this might be surgery for immediate closure. Or, the choices may come from the middle of
the spectrum, intermediate technological therapies meant to stimulate the
wound to heal, to force it onto the convergent attractor, including
pharmaceuticals, biologics, and physical and machine-based modalities. |
|
|
|
|
|
As a machine, as a non-linear system, as a system subject to
control and chaos, there is a final aspect of chaotic systems, including the
wound system that must be appreciated – thermodynamics. As we have seen through just a few
introductory examples or equations, non-linear & chaotic systems can have
stable attractors. A system tends to
dwell or return to that attractor, unless some major upheaval forces it onto
another attractor. Attractors can be
thought of partly as gravity wells or charge wells, a big planet or a small
ion that other objects want to orbit or fall into. Consider for example a planet with a
satellite. The satellite has kinetic
energy that keeps it in orbit.
Alternately, our friend the space alien might have some technology to
arrest the motion of the satellite to otherwise keep it suspended in a given
position, in which case the object has potential energy, ready to be
converted back to kinetic energy as soon as it is allowed to drop. If the object drops, or if its orbit
decays, then the system is giving up energy, and the orbit and objects are
converging. The space dude could also
turn on his thrusters, put energy into the system, and elevate the orbit via
increased kinetic energy, even to the point of escape velocity, i.e. divergence
between the objects. Between
convergence and divergence are orbits of variable period, depending on the
energy in the system. This is no
different than the attractors and dynamics of the Mandelbrot set. All objects have a certain basal energy. (Their mass per se is also energy, E = mc2,
but that is not convertible to anything else under common everyday
circumstances and need not be further considered.) They have an obligatory amount of energy
due to temperature and pressure. This
Kelvin energy cannot be converted to anything else without giving up heat. It can be transferred to other objects, but
if any of it is lost, then the temperature of the body diminishes. Closely allied to this basal energy is the
concept of entropy, the amount of heat gained or lost at a given
temperature. Entropy will always
increase in the “downhill” transfer of energy, so it reflects conditions of
equilibration or convergence. Objects
can also have “free energy”, additional energy beyond their Kelvin
temperature (Gibbs energy). It is
“free” because it is readily convertible without changing temperature or
entropy. This is kinetic energy and
potential energy, as stored in the separation of gravitational or charged
objects, or in the elastic deformations of a spring, or in the bonds of an
energetic molecule. For the orbiting
satellite, free energy is kinetic. If
it sheds free energy, its orbit decays.
If it gains energy, its orbit is elevated. If an elevator is stuck on the 5th
floor, it has a certain amount of free energy stored as gravitational
potential energy. If it goes up, more
free energy must be put in to elevate it.
If it descends, it sheds free energy.
Total system energy for an object is its enthalpy. Its obligatory basal energy due to heat and
pressure is its kelvin energy. The
difference is the free energy which is convertible and reusable. Consider an ideal spring or clock pendulum in motion, or a
satellite in orbit. The oscillation
rate of the spring and pendulum, and the height or period of the orbit will
depend on how much free energy is in the system. However, if it is an ideal perfectly
elastic or conserved system, then it oscillates or remains at altitude without
needing new energy (except to replenish whatever little is lost “internally”
due to friction and the like, e.g. the clock pendulum needs a wound spring or
weights). Nature and engineered
systems are full of such systems where, except to replace internal losses, no
other energy is needed to maintain motion or current state. Motion continues ad infinitum, because no
free energy is dissipated or transferred to another object. But if free energy is added to or removed
from the system, then system dynamics change, such as oscillation rate or
height or period of an orbit – the orbit goes higher or the clock ticks
slower. However, although certain
dynamics in the system are free energy dependent, others are not. If one records the motion of the
system, complex forms or morphology
can arise, passively and without the expenditure of energy. A Lissajous figure forms from a swinging
pendulum without requiring energy, and a clockface sweeps out overlapping
harmonic circles. Now, consider the
grass-sheep system. (This is discussed
as part of population logistics on slide 23.
You might want to read the next few slides, 21-22-23, then come back
here to finish this slide.) Grass mass
is converted to sheep mass via magic sheep enzymes, and sheep mass is
converted back to grass mass by magic fertilizer made by the sheep. It is a closed system of fixed mass that
flips back and forth between different levels of grass or sheep. It is also closed in that no energy is
transferred outside or to other entities, and thus no new energy is needed to
run the system other than some sunlight to refresh internal losses. The system is inter-convertible between
grass and sheep just by letting the system follow its own rules, letting it
“oscillate” so to speak between grass and sheep. Of course, the system does not really
oscillate – that would be periodic or harmonic motion – very linear or
non-chaotic. The whole point of the
non-linear logistics map was to show that the sheep-grass partition will vary
in complex ways that need to be iterated to see the moment-to-moment
values. So, it does not oscillate, but
it does go “back and forth” in its own way, which could be construed as an
elastic interaction of sorts, or an orbit of sorts (which is why we refer to
those attractors as orbits). Even
though chaotic non-linear systems move in ways that are not harmonic and
cannot be explained by Newtonian calculus (such as by a spring or
gravitational equation), nonetheless the same thermodynamic principles
apply. When a chaotic system is in a
stable orbit, it continues on that chaotic attractor in the same way that an
ideal spring (without internal energy dissipation) stays in its harmonic
oscillation without needing new energy inputs. It is easy to understand that if I want to
boost satellite orbit or spring oscillation, I need to put free energy in, by
thrusters or stretching. For chaotic
systems, changes of state and transitions between attractors can be increased
by putting force or perturbation in from outside stressors, thereby adding
energy to the system. And just like
for a Lissajous figure, chaotic attractors can also have complex morphologies
that may not be energy-dependent (e.g. the bifurcation maps, cobweb diagrams,
and “mandeloids” seen on slides 14 & 23).
These patterns and morphological complexities are more a matter of the
feedbacks and controls in the system, more a result their intrinsic or
internal elements, rules, and dependencies.
These are the “wave shapers” in the system, rather than the free
energy, a matter of how the free energy is routed within the system rather
than how much is applied to or withdrawn from the system. As an example, I could regularly add an
unnatural new fertilizer to the grassy field, something that promotes grass
growth, but also makes it unpalatable to some of the sheep. This is a perturbation from without (equivalent
to adding or subtracting energy) which will markedly affect the dynamics of
the system, fundamentally altering the grass-sheep balance and shifting the chaotic
attractor of the system. Alternately,
I could just change the time of day that the sheep are unpenned and allowed
to feed. This change in the internal
rules (equivalent to neutral energy redistribution only) might very well
change some of the precise iteration-by-iteration values of the system, but
the general forms and morphologies of the attractor, and especially its basins,
moments, and “centers of gravity” are unlikely to change. In chaotic systems, including the wound and any other complex or
non-linear physiology, the system will move from one attractor or orbit to
another, or converge or diverge.
Trans-orbit or trans-attractor shifts, or convergence or divergence,
are equivalent to adding energy to the system, or taking it out. When a system is stable on an attractor, it
has no net gain in energy or information.
Thus, attractors are like basins of stability, and the system will
break away only if enough energy is put in to elevate it above
threshold. The concept of basins or
wells of energy stability are of course common in nature. Consider certain exothermic chemical
reactions or a nuclear fusion or an action potential in a nerve or muscle
cell. An activation energy is
required, something to boost the system to its threshold or trigger point,
and then reaction is autonomous, releasing large amounts of free energy from
broken bonds. The system has to go
“over a hump”, taking a bit of added energy to climb over, then yielding much
larger amounts as it slides downhill, just as if it was falling in a
gravitational field and giving up potential energy. Of course, once the system has settled into
its new basin, there is no going back, at least not easily. If the reaction is nominally reversible,
all of the dissipated energy has to be put back in again to boost it up over
the now much higher hump. When chaotic
systems hit stable attractors, it is as though they have gone over such an
exothermic hump, giving up free energy, finding it “easier” to dwell where it
is rather than move to another state or attractor. Energy inputs by extrinsic stressors are what
can elevate the orbit again, and when the stressors are relieved, the system
can settle back into whatever attractor or basin it can “thermodynamically” –
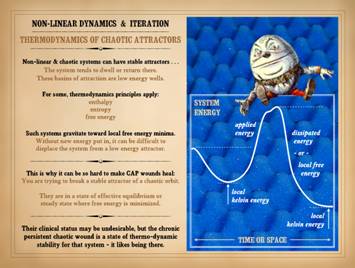
or just plain “dynamically” – occupy for the current conditions. Chaotic systems are generally not in equilibrium nor steady
state, but it can seem that way, or it can be defined that way. When a logistic map settles into a
bifurcated 2-value state (i.e. for A=3,25 on slide 23), that certainly could
be a valid instance of equilibrium.
Any n-value orbit that regularly returns to its multiple values could
also be defined that way if you choose.
Of course, for a long-orbit attractor, you may never see the return to
base value, and not ever for a converging or diverging system. For chaotic non-linear systems, the general
dynamics are different than equilibrium or steady state, but they can be
loosely associated for certain circumstances.
Low energy wells and stable basins of attraction will seem steady or
equilibrated when the system refuses to budge – like for a chronic wound that
never really changes regardless what you do.
When a chaotic system “gets stuck” in a low energy basin, it has
effectively undergone an exothermic reaction, giving up free energy, dwelling
at a point of low kelvin or basal energy.
These basins are local minima, either in time or space: (1) time, the
system may dwell there for a certain time, then applied energy or
perturbations break that cycle and lift the orbit, or (2) space, in a complex
system like the wound, there are zones and strata and fluxes that may make
one area be in a state different than a neighbor. When the system has found a stable
attractor or energy well, it can be difficult to displace it. This is why it can be so hard to make chronic
and pathological CAP wounds heal. You
are trying to break a stable attractor of a chaotic orbit. The system, the impaired wound, is in a
state of dynamical stability where free energy is minimized, and displacement
from that basin of attraction requires new energy or perturbation from the
outside. Obviously, if the system with
its own closed set of elements or actors has found a stable dynamic and wants
to dwell there on a closed repetitive orbit, it just simply isn’t going to go
to another orbit on its own. Some
player from the outside must come in and persuade or force it to do something
different. Those forceful outside
players are the deliberate therapeutic interventions that are prescribed and
administered for the sake of trying to cure the wound. For auto-immunopathic ulcers, where the
wound has become “intrinsified”, simple dressings, hygienic care, and basic
wound healing dependent surgical repairs are not sufficiently forceful to
raise the orbit or break the attractor.
The forceful interventions which can change attractors are the
pharmaceuticals, devices, and other modalities which suppress the primary
disease, used in partnership with potent technology based wound treatments
such as stimulatory and regenerative therapies. It is important to understand that for
chronic pathological wounds, their clinical status may be undesirable, but
the chronic persistent chaotic wound is a state of hard-to-break dynamical
and thermodynamical stability for that system. Humpty Dumpty, illustration by English
illustrator Sir John Tenniel (1820 –
1914), for Through the Looking Glass by Lewis Carroll (chapter vi). Illustrating the concept that “you can’t
make a chicken from chicken salad”, Humpty’s great fall epitomizes the
difficulties of sliding down an energy well – falling into a stable attractor
– and not being able to climb out so easily, not even with all the king’s
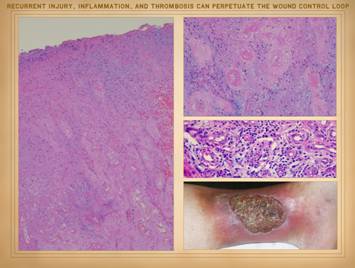
horses and all the king’s men. |
|
|
|
|
|
It should be clear by now that dynamical or thermodynamic
stability may be in synch with or at odds with clinical desirability, but that
a sick ulcerating wound is nonetheless a stable attractor. This raises the inevitable question, why then
does the CAP wound get into a state of dynamical stability even if that state
is counter-productive to health? If
the control loop is supposed to work toward restoring a reference, why does
it become complacent about an unconverged status? This has already been answered in part – it
is inherent in the principles of non-linearity, chaos, and attractors. At this point though we need to acknowledge
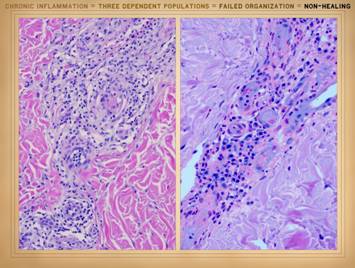
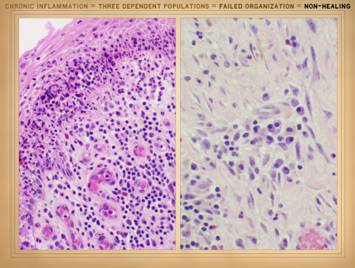
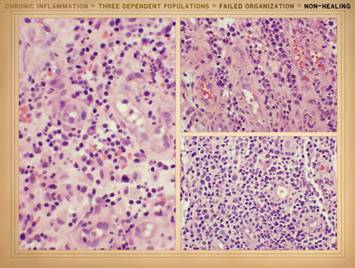
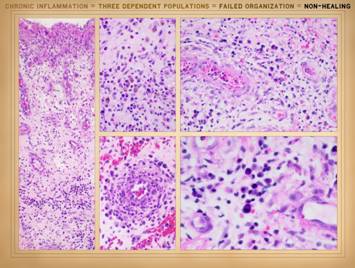
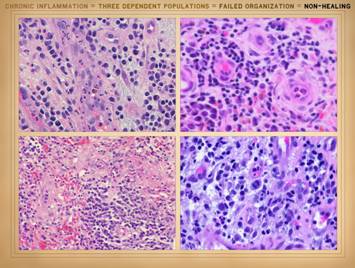
the actual physical structure or biological components of the wound. It is a set of cells. In Part 2 we saw that when wounds become
auto-immunized and pathological, that the normal two cell populations of the
healthy wound, acute inflammation and wound module, are joined by a third
population, chronic inflammation. As
we discussed on slides 2 & 3 of this section, the wound as an organ is
defined by its cell populations. To
understand how these populations interact and either succeed or fail to
restore a stroma, we must now look at another aspect of non-linear dynamics:
population logistics. |
|
|
|
|
|
Understanding population dynamics starts with understanding its
simplest scenario, a single population whose growth is limited by available
resources. First, think about
population growth intuitively. You
start with a small number. They
reproduce. Their descendents
reproduce. If within a given time interval
there will be a consistent fraction of the population which spawns, then in
each time slice population increases by that percentage. For example if population is P and
growth rate is r, then Pt1 = rPt0. After n iterations, Ptn
= rnPt0. This
obviously is a form of exponential growth.
To be more formal and continuous, dP/dt = rP, the solution of
which is simply Pt = Poert. The problem is that in the real world,
there are few opportunities for a population to undergo totally unconstrained
exponential growth. Resources will be
limited – space, food, whatever – and this will slow growth until the
population’s resource utilization matches the resource supply rate, or until
it has occupied the available space.
The simple concept of exponential growth has to be modified to account
for resource limitations. The subject
is called population logistics, and in its simplest form, it derives from the
Verhulst equation. The Verhulst equation (aka logistics equation) concerns a
population in which space or substrate supply is limited, meaning that the
population can grow only to a maximum.
If substrate supply remains constant after achieving the maximum
population, then the population stays there, new individuals replacing those
that are lost, a steady state. If
substrate supply rate starts to dwindle, then the population will start to
decline, the curves going down in reverse of the uprise. Let us focus on the uprise. The curve starts out quickly, because a
small population has a lot of space or resource to grow into. As the space gets more crowded, there are
more citizens and fewer free resources, so growth rate must slow. Eventually population caps out at the
maximum sustainable. Intuitively, the
curve should be sigmoidal or plateau-like.
That is in fact what happens. Start
by defining P = actual population, K = maximum permissible
population (“capacity”), and r = maximum possible growth rate (the
intrinsic reproduction rate if there were no resource constraints). The population will grow according to dP/dt
= r P (1 - P/K). This means that
the change in population per time is proportional to three parameters: (1)
the existing population P (the basic predicate of unconstrained
growth); (2) the space available for new population to grow into (1 – P/K)
(remember, P/K is a fraction less than 1); (3) the intrinsic growth
coefficient r. [ The analytical
solution of this equation is a simple integration. The trick to doing this integration is to
do a preliminary transform on the equation.
Start with dP/dt = r P (1 - P/K). Next, divide both sides by K: dP/Kdt = (1 - P/K) rP/K. Now, define x = P/K (this value is
the population fraction, the portion of total capacity that is currently
populated). The equation is now dx/dt
= rx(1-x), which can be integrated easily. ] Notice the dynamics of this equation. When x is small (early population), then
(1-x) is nearly 1, and the growth rate dx/dt is close to rx, the unconstrained
maximum rate. As population matures,
(1-x) governs the values, and as x nears 1 (available space nearly full),
then (1-x) approaches 0, and growth ceases.
The slide shows the Verhulst equation graphed for various values of r. The Verhulst equation as just presented is linear – i.e.
continuous, differentiable, and a true function of a dependent value
(population) mapped one-to-one onto an independent variable (time). It tells you the population as a function
of time. It is orderly, because it has
no dependencies other than time. The
key stipulation for using this equation is that the growth system has one
population that is resource limited.
As shown in the next slide, the dynamics get more complex as multiple
populations appear and compete with or otherwise influence each other. |
|
|
|
|
|
The Verhulst equation describes intrinsic population dynamics,
but the way it applies or behaves changes as the system gets more complex. Population levels and behavior become very interesting
when additional populations, population dependencies, and resource
restrictions are put into the system.
To illustrate, we will start with a simple scenario. Consider a big field full of grass with
some sheep. The sheep eat the grass,
grow strong and fecund, and beget more sheep.
If the grass maintains a fixed mass year by year, then there will be a
maximum number of sheep that can be supported, and the sheep will grow into
this fixed resource “space” according to simple Verhulst dynamics. What happens though when the resource
supply rate is not fixed, in fact it competes with the sheep? How?
Let us “close” the system so that there is a fixed biomass divided
between sheep and grass. This means that the grass mass decreases as sheep
mass increases. Eventually, the sheep
outgrow the available grass. The sheep
population will then start to decline, and as it does, grass mass again
increases. In this scenario, the grass
is not supplied at a fixed rate. The
grass is an active population itself, just like the sheep, and it can be
depleted but then it can rebound.
Sheep and grass are two mutually interlocked populations, at times
supportive, at times confrontational.
Nutrition, starvation, predation, cultivation are the intertwined
dynamics. The problem gets even
hairier if you then throw in a third element such as wolves to eat the
sheep. In this scenario, can we
calculate the population of sheep or grass as a matter of time? Not so easily. The Verhulst logistical equation still
applies in principle, but now it must be applied to two populations, and
those two are contingent on the other!
The balance between two interactive populations cannot be calculated
by a continuous linear equation, because this has now become a “non-linear”
problem, the heart and soul of real world complex systems. There is no way to calculate, as a direct
analytical function, how many sheep or how much grass is there, neither one
as a function of the other [ sheep = f (grass) ], nor parametrically in time
[ grass = f (t); sheep = f (t) ].
Nonetheless, this problem can be solved, but as for any non-linear
complex system (aka non-linear dynamics) the problem is solved by recursive
iterations, using the iterative form of the Verhulst equation, the “logistic
difference equation” xn+1 ! Axn(1-xn). Notice that it is still the logistical
equation, with x being sheep, and (1-x) still being the space available for
sheep to grow into. However, (1-x) is
also grass, and grass is an independent population, and from the grass point
of view, grass is the dominant variable g, and sheep are just the available
resource (1-g). Growth rate A is also
present. It turns out that the dynamics of this system, the amount of
sheep or grass, has a very strange and wonderful set of back-and-forth
numbers iteration by iteration or year by year. The table shows iterates for various values
of A. (Note that this table was
created on a spreadsheet. For anyone
interested in this subject, these equations can be experimented with using
tools that most likely are already on your desktop.) When iterated, the value of x will do 1 of
4 things (depending on the value of A):
converge to zero, diverge to
infinity, enter an N-period orbit, never settle (chaos). See how this is similar to the behavior of
the Mandelbrot? This is the general
behavior of non-linear systems. Notice
that at lower values of A, the iterates will wander through a few values,
then converge to a fixed value, e.g. for A=1.50, x stabilizes on 0.333 (a
1-period orbit). As A values increase,
it takes more and more iterations to converge to a fixed value. After that, the values bifurcate, flipping
back and forth between 2 values, e.g. x =
(0.496, 0.812) for A = 3.25. At
yet higher A values, the orbit bifurcates again. Amazing, but that is the way it really
works. The bifurcation diagrams in the
center map the orbit values versus A.
What is obvious is that a simple iteration like this can lead to intricate
complexity and seeming mayhem, i.e. mathematical chaos. On the right are cobweb diagrams that map
the sequential values of the iteration on a graph that has the linear form of
the equation (parabola) and the identity line. These show that although values may jump
around erratically on a large orbit, that nonetheless they stay bound to a
well or basin or attractor or compact state space. Population dynamics are deterministic and
chaotic, but not random. Compare this
to the boat captain and his alien observer (slide 13). If you know all details and parameters of
the system, you can calculate the next iterate. If you are not LaPlace’s Demon and do not
know all relevant information, you can still observe the general attractor of
the system and the limits of its allowable values, but you cannot be certain
what the precise next value will be. Now, consider that these two complexly interdependent
populations, grass and sheep, were spatially separated at the start. Suppose that the first few sheep were
confined to just a corner of the field, and that they stayed in their corral
because there was more than enough grass in that corner to meet their
needs. However, as the population
increases, then in their quest for food, the population will migrate outward
to greener areas. Whether by random
wanderings (aka random walks or diffusion) or by taxis and tropism
(explicitly drawn to the greener pastures), the sheep eventually spread
through the field. If they were meant
to stay separated when sheep population was small, that sequestration falls
apart as time, population growth, and diffusion progress. Now, compare this principle of diffusion
and admixture to the events that occur in the wound, when inflammation is
sustained, and the inflammation and repair populations get mixed in time and
place. Grass-sheep or wolf-grass-sheep are not meant to be strictly
analogous to the wound and its 2 or 3 populations. For instance, the wound is not a fixed
biomass system as the grassy field was.
Relationships might seem reversed or counter-intuitive, e.g. lymphoid
induction of acute inflammation is bad clinically, but it is a
promotion-cultivation event between these populations. Nonetheless, for both of these systems, the
core principles of non-linear logistics apply. For acute inflammation, wound module, and
lymphoid inflammation, their appearance, uprise, then decay follow Verhulst
dynamics. When their intermix dynamics
become chaotic, then the value of any one of the populations varies in
complex ways. However, the chaos and
complexities merely reflect the dependencies or contingencies that develop and
feed back between these populations:
predation, deprivation (starvation), cultivation, sustentation
(nutrition). They can promote each
other, compete with each other, starve-attack-inhibit each other, cultivate or
nourish each other. The physical
realities of wound versus grass-sheep may be different, but their dynamical
physics are the same. The main message
is that in these complex systems of interacting populations, the dynamics can
be strange, seemingly unpredictable, and at times locked in to “attractors”,
states of being, from which it is hard to break away. The physics of non-linear dynamics explain
that this is the normal natural behavior of complex interactive populations. |
|
|
|
|
|
The application of population logistics to the wound was implied
in the last paragraph. This slide
looks at wound logistics for the healthy one-shot wound. Healthy physiologic wound healing is initiated by injury &
thrombosis, then conducted by two cell sets, acute inflammation & wound
module. It is a sequence of one-shot
self-completing linear events: primary injury & thrombosis, then acute
inflammation, then wound module. The
sigmoidal rise and fall of each population shouldn’t be surprising. These populations, acute thrombosis, acute
inflammation and wound module start from zero, no population. Their precursors are there, but the
activated species are not. Once
activated, they beget more of themselves.
Thrombosis begets more of itself by auto-amplification and recruitment
of more precursors. Acute inflammation
(leukocytes) beget more of themselves by recruitment. Angiocytes get more of themselves by
mitotic proliferation. For all, the
dynamics are sigmoidal, fast on the uprise when amplification is largely
proportional to extant population, then topping out as primary triggers or
inducers decay, equivalent to a reduction in a vital resource. The downslope occurs as triggers completely
disappear, a time lag or phase shift between the triggers and the response of
the population, but still sigmoidal-Verhulst. Each of these events or populations is an integrator function to
trigger the next population. As each
decays, it stops inducing the next, so then the next population starts to
decay as well. When the system is
healthy and injury is incidental or self-limited, then the reactive events are
non-competitive populations that barely overlap or “see” each other. Each compartment triggers the next, but
there is no feedback to sustain or re-trigger events. Because there are no feedbacks or sustentation,
then the overall dynamics avoid complexity, recursion, and instability. |
|
|
|
|
|
We now look at population logistics in the pathological
wound. Pathologic wound healing occurs
when sustained injury, population persistence, non-linear feedback, & abnormal
population dependencies sustain the activity of any or all of the primary compartments.
The persistence of the primary disease or injury can obviously
be a cause of sustained abnormalities.
If the disease is sustained, then the response of the subsequent
compartments is to get “smeared out”. They
will extend in time, and their populations will increase under the effects of
the continued induction or stimulation.
The compartments will get increasingly admixed or exposed to each
other as time goes by, both overlap in time, but also admixture in the strata
of the wound. If the problem was just
a matter of continued primary injury, then the outputs would still be
linear. The populations of acute
inflammation and wound repair could be calculated from (1) the transfer
function or relational equation that defines the response (dependent
variable) to the injury (independent variable), and from (2) the
deconvolution of the output that can separate values related to a “smeared
out” input. The whole system would be
sustained as long as injury is active, but the dynamics would be relatively
smooth, well-behaved, and related to the input. Inflammation would subside (Verhulst) and
then repair would complete itself (Verhulst) as soon as primary injury was relieved. The real problem is when feedbacks develop that “chaoticize” the
system. Remember the circular
relationship between thrombosis and inflammation, and their effects to make
new wound which can then further promote the system: injury -> thrombosis -> inflammation
-> injury -> etc. etc. etc. Once
sustained injury leads to sustained inflammation, then these cycles feed back
to stimulate their antecedent events.
Once this happens, then the system state or output loses all
functional relationship to the input – i.e. it is chaotic. The dynamics of this mix include a variety
of promotional and inhibitory effects between populations and
compartments. These are the “negative”
population dependencies which will tend to promote or sustain the pathological
state, making it harder for the system to settle back to its linear Verhulst
dynamics and thereby heal the wound. The
nominally normal 2-population wound (acute inflammation and wound module),
can therefore develop non-linearity, persistence, and chronicity if primary
injury or disease are sustained. As
discussed in detail in Part 2, “primary injury or disease” can be any
condition of repetitive or sustained trauma, inflammation, immunity, allergy,
infection, thrombosis or micro-occlusion, etc. These dynamics worsen when pathology, negative dependencies, and
chronicity eventually induce a 3rd population of cells, lymphoid
inflammation, aka “chronic inflammation”.
This has the effect of taking an already chaotic 2-body problem and
turning it into a 3-body problem, further complicating the mix of mutual
promotion, inhibition, and competition.
(This is like going from grass-sheep to grass-sheep-wolves. If one considers thrombosis to be its own
population in this mix, then it is a 4-body problem.) This complex mix of dynamics cannot land or
converge, consistent with the non-linear form of logistical dynamics (the
logistic difference equation) rather than the linear form (sigmoidal Verhulst). As we will see soon, this third population
is an autonomous negative influence on the system, acting as a sustained
primary injury. Even when initial
disease or injury is then eliminated, the effects of the third population are
comparable, maintaining the chaotic dynamics that are already taking
place. In the 3-population wound, perpetual
complexity arises in the logistics of:
injury & thrombosis -vs- acute inflammation -vs- wound module -vs-
chronic inflammation -vs- injury & thrombosis –etc.-etc.-etc. At this point, we can actually draw a closer parallel to the sheep-grass
analogy, by looking at the dynamical symmetries in this mix. Normally, acute inflammation (ai) has an
inducing or proliferative effect on the wound module (wm), ai gÉ
wm. In a normal wound, that
is all there is to it, acute inflammation turns on healing, and each phase
runs its own course. (Normal healing
does not have direct inhibitory feedback on inflammation. Instead, inflammation runs its course, and
if there is no further injury, then it extinguishes itself as repair ramps
up. To the extent though that repair
keeps the tissues in a “good state” that inflammation is not alerted to, then
the wound module can be seen as having some inhibitory feedback on acute
inflammation, ai Vf wm.)
In the chronic pathological wound, chronic inflammation (ci) joins the
mix. Chronic inflammation has an
effect to suppress or upset the wound module, disorganizing it or retarding
its kinetics, ci gV wm.
The altered wound module in turn is creating auto-sensitizers and
exposed antigens which fuel the chronic inflammation ci Éf
wm. Note the symmetries in
these dynamics: acute inflammation
begets wound module; chronic
inflammation suppresses wound module;
normal wound module suppresses acute inflammation; altered wound module promotes chronic
inflammation. Thus, we have a system
of mutual feedback, mutual promotion and inhibition, mutual predation and
deprivation, mutual induction and suppression. These are precisely the types of dynamics
that affect the simple grass-sheep system. There is a reason that chronic and pathological wounds act
“locked in”, now a bit better, now a bit worse, but fundamentally unaltered
over long periods. They are locked
into a dynamical attractor where acute inflammation, an altered wound module,
and chronic-immune-lymphocytic inflammation compete and promote and can not
easily escape. These dynamics are
easily understandable via the principles of non-linear population logistics. When healthy (the 2-population wound), recurrent primary injury
and thrombosis are needed to perpetuate the wound and the wound control loop. When intrinsic chronicity occurs (the 3-population wound), the
system is locked into a persistent chaotic attractor without needing new
inputs. |
|
|
|
|
|
This slide emphasizes the last few comments from the preceding
slide. When the wound is intrinsically healthy, with the normal and
expected 2-populations of acute inflammation and wound module, then recurrent
primary injury and thrombosis are needed to perpetuate the wound and the
wound control loop. When intrinsic
chronicity occurs, chronic inflammation creates the 3-population wound, and the
system is locked into a persistent chaotic attractor without needing new
inputs. Perpetual complexity arises in the logistics of: injury & thrombosis -vs- acute
inflammation -vs- wound module -vs- chronic inflammation -vs- injury &
thrombosis. These inter-mixed and
inter-dependent populations have abnormal dependencies and feedbacks leading
to multicontrol and n-body dynamics, loss of sigmoidal one-shot behavior, and
chaos and non-converging attractors.
As problematic as this might be from a clinical point of view, this
system can enter an attractor that is dynamically and thermodynamically
stable, a low energy well that it cannot so easily escape. On the left panel, the wound control loop is shown, with
extrinsic injury and thrombosis summed in (i.e. primary disease – trauma,
allergy, immunity, hypercoagulability, etc).
Obviously, if the loop was otherwise running as a one-shot, winding
down as the wound healed, then these added events would simply expand the
wound, cause more inflammation, and elevate the whole system to an earlier
more active phase. On the right,
chronic inflammation has been added in with the other major dynamical
compartments (injury & thrombosis, acute inflammation, wound
module). This adds a variety of new
dependencies, both promotional and inhibitory, which complexify the patterns
of feedback and thereby lead to chaotic behaviors. |
|
|
|
|
|
The top picture shows a young wound, still open, going
through the process of active proliferative wound healing. This is a healthy wound. The architecture of the wound module is
present, but in any given area, the structures are largely amorphous or loosely
organized. In the bottom
picture, an old scar has remodeled itself back to nearly normal fascia. The arrangement of angiocytes and
fibroblasts, vessels and connective proteins, is very nicely ordered and
periodic. There is no blueprint of how
to do this, yet it happens dependably well in the healthy patient and
wound. The Main Control Loop drives
wound healing toward the system reference.
How does it know what the reference is? How do wound module elements know to
organize into the required form that satisfies the reference? To study the physics of wound healing and wound pathology, we
have so far looked at three foundational subjects in non-linear
dynamics: (1) control, (2) chaos,
attractors, n-body dynamics, (3) population logistics. The answer to the above question comes from
a fourth subject in non-linear dynamics:
cellular automata & self-organization. |
|
|
|
|
|
The whole wound healing system represents another foundational
concept in non-linear dynamics – it is a class of cellular automata. Self-organizing
automata are systems in which individual elements, “cells”, have a set of
strict deterministic rules governing their behavior and how they must
interact and evolve with adjacent or local cells. If you throw them all into a pot, they will
sort themselves out, generally ending up with complex highly organized
structures based on just a few simple instructions or relationships. Biological systems which are undergoing embryogenesis, growth,
and repair are all self-organizing automata.
Growth and repair have no architectural blueprints to follow. Cells and chemicals just do what they do,
obeying basic physical, chemical, and biological principles (the
deterministic “rules” of the system).
In so doing, proper structure develops. Dynamical and thermodynamical principles of
energy and conservation ensure that systems will passively and blindly “seek
the path of least resistance”, undergoing reactions or transformations that
tend to dissipate free energy and maximize entropy and thereby seek zones of
energetic or dynamical stability.
These thermodynamic principles of efficient assembly are epitomized in
the term “parsimonious self-organization”.
This principle states that automatically organizing systems will take
the route to self–assembly that takes the least information or energy or
effort. Essentially all organized structures and patterns in nature
qualify as self-organizing automata, not just biological systems. In other systems, “cells” are the
functional or interactive units that constitutes the base scale of the
system. They may be physical units, or
they may be abstract partitions or tilings of a space or medium. The key premise of cellular automata is
that the behavior of each cell is determined locally by its interactions with
its neighbors. Some behavior or change
in state is induced as a reaction to what the neighbor is doing. Systems can be defined to have
“neurological” functions (orders are issued from a central command) or
“endocrine” functions (status can be telegraphed over a distance). However, the reality is that an extraordinary
number of systems in nature and biology are just local “cellular”
interactions. In biological systems,
the dynamical “cells” are likely to be real biological cells. In the wound, each cell type or population
has its own genomic program and “rules of engagement” that specify how it is
to react to other cells or environmental features, and likewise how it is to
influence the others. These are the
rules of the system, and they govern what type of structure occurs. The illustrated examples on this and the
next slide should give a sense of what cellular automata are and how they
operate. Left top: these are
two examples of abstract automata generated by computer. Each pixel in the image is assigned a
random value, black or white. On each
iteration, the state-value-color of the pixel is tested and altered based on
a strict set of rules. The rules
typically are something like “if three neighbors are black then become white,
else no change”, or “if the guy to the right and the guy below are black then
become black, else become white”. I
cannot recall what rules I used to generate the these two outputs, but that
is irrelevant. What happens is that as
the system is iterated, beautifully organized patterns show up and then
remain stable (no changes on subsequent iterations) – just like a point on
the Mandelbrot set or logistics map when it enters a stable orbit or
convergent value after so many iterations. Left center: Diffusion
limited aggregation (DLA) systems are a great example of a simple cellular system
that models many real world structures.
The assumption is that the central structure grows by the aggregation
of new material, and that the arrival rate of new material from the
peripheral environment is much slower than the aggregation rate onto the
structure. The system is iterated by
starting a “particle” at a boundary pixel, then letting it do a random walk
until it hits the existing structure, which is where it then sticks. This concept is extremely simple to model
digitally, and the left panel shows a computer generated output. On the right is a picture of manganese
dendrites on a desert rock, a nice example of the kinds of natural structures
that are formed by DLA dynamics. Below: These are samples of the
VT (Vascular neT) model of angiogenesis.
This model recreates the biological rules of angiogenesis based on the
diffusion of oxygen and angiogenic factors, and the sprouting of new vessels
on threshold conditions based on cell-to-network distances. The four panels to the left show the generation
of new vessels over four iterations as the host tissue grows (the four panels
have been resized to a common display size).
The two panels to the right match a VT output to a picture of real
blood vessels. The model accurately
recreates vascular morphology based on a few parsimonious rules applied
locally by repetitive iteration – i.e. a typical non-linear cellular automaton
based explicitly on the natural rules of the system. (See the Arimedica website for more
information about the VT model and angiogenesis: “Developmental Angiogenesis and the
Biophysics of Vascular Network Formation”, May, 2006.
http://www.arimedica.com/content/arimedica_vt_(slides)_2006-0516.pdf) |
|
|
|
|
|
Here are more examples of automatic self-assembly in biological
systems. They are easy to find,
because the body and its components have no means of organization other than
cellular self-assembly, parsimoniously organized. Remember, genes encode proteins, either
structural proteins or chemical process regulators (enzymes). There are no morphology genes, They do not male plans or drawings. It is in their inter-operations that
complex organization and macro-structure occurs. Simple chemical and physical interactions
establish the small sets of deterministic “instructions” which govern how
cells interact with local neighbors.
The self-assembly of tissues and organs then occurs
automatically. Non-linearity,
feedback, and control are implicit & explicit. Left top: The visual
cortex of the brain is organized into ocular dominance columns. The inputs from the right and left eyes are
separated into interdigitated bands.
How this separation occurs, and how these bands or “columns” develop
is a cellular automaton based on just a few rules. Local competitive promotion and inhibition
between populations is the crucial physics that governs the interaction. It is easy to model digitally by iterative
non-linear implementations of the rules.
On the left is a picture of real columns. The other two images are computer
generated. (1) Carreira-Perpin MA et al. A computational model for the
development of multiple maps in primary visual cortex. Cerebral Cortex, 2005,
15-8:1222-1233. (2) Miller KD
et al. Ocular dominance column
development: analysis and simulation. Science, 1989, 111:123-145. Left second: Animal coat
patterns occur as the result of the diffusion of local promoters and
inhibitors. Chemical diffusion is the
constitutive physics that governs the interactions. These patterns can be modeled easily on a
plane by iterating reaction-diffusion equations that let neighboring pixels or
cells decide if they turn or not to produce pigment. Murray JD. Mathematical Biology, 2nd
edition, Springer, 1993. Left third: The same
reaction-diffusion equations can model the on-off pigment patterns that
develop in mollusc shells. It is
analogous to a printer in which the shell is a piece of paper. The pattern is printed one raster at a
time, with each “character” being an automatic “cell” (and a biological cell
as well) that responds to the effects of its neighbors. Meinhardt H. and Klingler M. A model for
pattern formation on the shells of molluscs. J Theor Biol, 1987, 126:63-69. Below: Here are more examples
of how the VT model can recreate branched network structures. Remember, the VT model operates by testing
local threshold conditions of the distance between each cell and the network,
a good example of automatic “cellular” self-assembly. Left, bile ducts form under the same
dynamics as vessels, and they have the same patterns. Middle, the gastrovascular cavity of a
platyhelminth flatworm is easily modeled by VT, since it is the same
structure as our own vessels, governed by the same genes (vegf), occurring in
a species that phylogenetically has not yet separated the food accumulating
gastric system from the distribution functions of the vascular system. Right, the re-creation of veins in a plant
leaf. Wound repair and the wound module are just another example of
this kind of automatic self-assembly and parsimonious self-organization /
self-re-organization. When allowed to
function properly, stromal rebuilding is automatic and correct, without any
blueprint or collective knowledge by the cells of what is being built. |
|
|
|
|
|
The wound, i.e. the reparative events that mend the injured
tissues, is simply a matter of the stroma reorganizing itself. It is a process of self-assembly by the few
types of biological cells which constitute the system. The rules or interactions which govern the
process are functions of the interactions between nearby cells. As such, the wound is a cellular
self-organizing automatic system – a cellular automaton – made of real
biological cells. Their deterministic
interactions are epitomized in the Main Control Loop. The photos are portraits of cells that participate in the
healthy and the pathological wound. Each
normal intrinsic cell - monocyte, angiocyte, fibroblast, keratinocyte - has
an assigned job. If allowed to
function properly, stromal rebuilding & wound closure are automatic and
correct. When the wound or its cells
are pathological, than self-organization is disturbed or fails, and wound
healing is disrupted – because wound healing is nothing more than the
self-organization of these cells.
Problems happen when chronic inflammatory cells appear. They disturb the orderly function and
organization of the other cells that are meant to be there, arresting or
disrupting their intended organization. The organization of the wound is an automatic event when all is
healthy. Recall from the images on
slide 27 that the goal of all of this organizing is to get to a stable
form. Those forms (mature normal
dermis and fascia) tend to be highly structured and periodic, more
“crystalline” than amorphous or glassy.
In parsimonious automata, they get to that form with minimum
information or energy, and that is the beauty of these systems, that such
dependable and consistent form arises from so few inputs or
instructions. If you look at the top
pair of examples on slide 28, the computational automata, the fields start as
random white noise. As the system is
iterated, they organize toward the forms shown, and once they reach the final
output, it no longer changes. Once it
reaches the form shown, each subsequent iteration produces exactly the same pattern
– it has become totally stable. This
is like the non-linear logistics equation that gets to a stable single value
or orbit after a few iterations. It is
like the Mandelbrot set that likewise converges to stable orbits after so
many iterates. Do you see the common dynamics
of these non-linear systems? When
convergent, they will stabilize to a dependable extremely stable form just by
following the simple rules. Even if the
resulting morphology appears very complex, it is nonetheless structured from
a very minimum set of rules and is ultra-orderly, and it is ultra-stable if
the system was able to fully converge.
Once a wound is fully re-organized back to final
scar-stroma-dermis-fascia, once it is fully converged, it is stable. Angiocytes and fibroblasts, vessels and
connective fibers are then settled into their final positions and forms that
will remain essentially unaltered for the remainder of the host’s life. (Whatever remodeling subsequently takes
place in the stroma is a long time-base or slow decay-rate event that is then
part of normal basal biology, not part of the original automatic assembly of
wound healing.) When the wound is pathological and organization is disrupted,
the system is still a group of cellular automata. The problem is that it cannot converge on
the intended form. It remains in a
loose state of quasi or unsettled organization, trying to organize but
remaining mixed up, remaining glassy and amorphous rather than crystalline,
fluid or flowable rather than solid, plastic rather than elastic. As “iterations” or time continue, the
structure can keep reorganizing and re-morphing, with cells and structures
appearing, disappearing, flowing, shifting, reshuffling. It will never be fully settled until
adverse “chaoticizing” dynamics that elevate the orbit are controlled and the
system is allowed to converge. The
failure to converge or fully organize is simply a consequence of non-linear
dynamics: an iterative or n-body or
multi-population system getting stuck on an attractor that cannot converge or
settle. For the wound, breaking it out
of a high-orbit attractor means controlling applied perpetuators such as
continued disease, injury, or thrombosis, or eliminating the intrinsic
pathologies that develop due to abnormal populations and population
dependencies. Recall too that the wound is a closed loop control system. How does that relate to automata? It has been implied so far that these
systems all function rigidly with strict algorithmic repeatability. That is certainly true for the
computational automata shown on slide 28, but it is not true for biological
systems. Biological systems such as
the wound are highly deterministic, but they must interact with the world
around themselves, and that world is going to throw a lot of unexpected
stresses, perturbations, and random variability in their way. Without control, the final output (a
morphological structure in the case of the wound) would have to be
pre-programmed, aka “calibrated”, and the environmental factors would throw
that calibration off. With good
control, healthy cells will react properly to whatever unexpected or aberrant
conditions are present. Determinism
and control will prevail, and the structure or morphology will form correctly
in spite of the environmental stresses.
That is explicitly the value of closed loop control over strict linear
open loop programmatic determinism, that errors and stresses can be absorbed,
overlooked, corrected, and accommodated.
Control makes self-assembling biological systems robust and error-tolerant,
unless the system controllers are sick or else the stresses on the system are
so severe as to overwhelm its inherent capacity to react and correct. The “dysdynamia” we are talking about should be distinguished
from conventional disease. From the
clinical perspective, the non-healing wound is a “disease”, a morbid state
that interferes with health or function.
However, the intrinsically pathological wound is not a divergent wound,
not ulcerating and getting worse. It
just is not healing. In the chaotic
wound, what is sick is just the collective interactions of otherwise healthy
cells doing what they are programmed to do in response to local stimuli. What is disrupted is ONLY the aggregate
organization and self-assembly. In the
chronic chaotic intrinsically pathological wound, not one of the cells shown
is intrinsically pathological. Each
cell and cell type is intrinsically correct or healthy, trying to do what it
is meant to do. If it is failing, it
is because it is being besieged, restrained, or deprived by another of these
cells. If it is abnormally overactive
and thereby disrupting things, it is because something else is stimulating,
up-regulating, or otherwise turning it on.
None of these cells is sick.
Just their collective interactions are altered. From a clinical point of view, the system
as a whole is sick. From a physics
point of view, this is just the expected dynamically and thermodynamically
mandated behavior of complex multi-control systems. Dysdynamia of the system is the intrinsic disease of wound
healing. Would it not be correct
though to say that genetic or metabolic diseases of angiocytes or fibroblasts
are themselves intrinsic diseases of this system? Yes, in principle. In actuality though, there are few if any such
diseases. That may sound strange,
given the spectrum and nature of other diseases that afflict the human
condition. Nonetheless, there are few
intrinsic disorders of the stroma and wound repair cells. For an explanation, see the addendum after
slide 34. |
|
|
|
|
|
At the start of this series (Part 1, slide 2) we saw examples of
wounds that had no net change over long periods. Convergent wounds which are healing are the
clinically desirable state. At the
opposite end of the spectrum is the divergent sick actively pathological
wound. While this is the clinically
adverse state, it is dynamically easy to understand, and the principles of
treatment to control disease and active ulceration are usually easy to
implement. In between are the
non-convergent chaotic wounds, the source of prolonged frustration, even
exasperation. These wounds refuse to
heal or even make progress, in spite of numerous reasonable treatments, even
when gross pathology & causative disease are controlled and acute
ulceration and inflammation are subsided. We stated at the beginning that there is a reason: that these wounds go back and forth but get
no better; that they cannot
spontaneously climb out of this attractor;
that multiple therapeutics are often of no benefit; that adverse behavior is independent of the
primary pathology. Furthermore, these
reasons cannot be understood: by
looking at any individual cell or chemical or gene; by analysis of any dependent-vs-independent
experiment; by any “conventional
bioscience” type of experiment; by any
type of randomized controlled trial. We are now in a position to understand why this is all so. Wound failure is not the result of a gene
mutated, and it is not the result of a chemical or metabolic imbalance. It is not the result of chronic
degenerative changes in a specific organ or tissue, nor the result of
obstruction, perforation, or other anatomical disruption. It is the result of a bunch of cells not
being able to properly inter-operate and self-organize. The science required to understand this is
not biochemistry and cell biology, not genomics and proteomics, not classical
anatomy and physiology. True, these
disciplines apply to understanding individual components of the wound, but the
science required to understand how wound elements inter-operate and organize
is physics. Specifically, the
applicable physics for wound healing is non-linear dynamics. So far, we have looked at this subject from
the point of view of several major aspects:
control, chaos, populations, and automata. These aspects will now be brought together
for an integrated view of the physics of wound healing and wound failure. |
|
|
|
|
|
In our study of non-linear dynamics and the wound, we began by
showing that normal wound healing is a regulated process, a feedback
controlled loop that senses the state of the wound and can drive the wound
healing machinery to restore an intact epithelialized stroma. When the control loop operates without
perturbation or repetitive re-injury, then the healing of incidental injuries
and wounds is a one-shot response, with reactive then reparative components
ramping up then decaying as they complete their appointed tasks. In fact, each major block or component of
the process – thrombosis, acute inflammation, wound module and repair – has a
similar one-shot profile, each triggering the next into existence, each
extinguishing after its task is accomplished. Problems begin when some sort of primary injury or stressor on
the system is sustained. There is a
crucial physiological interaction between inflammation and thrombosis, each
causing the other, and each able to be triggered by various primary
conditions of coagulopathy, immunopathy, angiopathy, panniculopathy. Necrosis and ulceration are caught in the
middle of this complex set of interactions.
Repetitive or persistent or primary pathology can trigger the
thrombosis-inflammation loop into a sustained state. This condition feeds back into the major
events of the primary sequence of wound reaction and repair. Once thrombosis and inflammation are
sustained, the one-shot profiles of healthy response and repair are
destroyed. There are now multiple
events, populations, control blocks, whatever – multiple “bodies”, i.e.
N-bodies – feeding back in a system of complex multi-control. Each element or component is responding
exactly as programmed. In fact, if you
were to look at small slices of the system, you would continue to see proper
linear or exponential or sigmoidal responses to incidental changes over short
time scales. The problem is that the
aggregate whole has become disarrayed.
The system remains deterministic, operating according to its rules,
but the overall behavior is one of back-and-forth, never crossing the finish
line, in patterns that defy representation by conventional geometry or
algebra, i.e. chaos. Once the
aggregate wound response-repair system is in a chaotic state, the tail end of
wound module and repair cannot run its course and succeed at its own
business, which is the reorganization and self-assembly of the stroma. |
|
|
|
|
|
Once we understand that the wound is a feedback regulated
control system, that lays the foundations for understanding that chaotic
dynamics can occur. Those chaotic
dynamics are realized when abnormal perturbations, external forces,
retriggers, and continued disease and injury keep boosting the system
components into sustained states or activities such that one-shot dynamics
and settling are preempted or disorganized.
The problem is especially disorganized when multiple elements with
multiple interactions (n-bodies) participate in the chaos. If all we talk about is chaotic systems in
the abstract, that suffices to explain wound failure. However, these principles become more
tangible when we start to address the physical reality of the wound, a set of
self-organizing cell populations. The
interactions of these cell populations under healthy circumstances is the
basis for normal one-shot responses, based on the principles of Verhulst
sigmoidal population logistics. One of
the well founded principles of linear dynamics is that when constraints and
feedbacks are placed on mutually contingent populations, chaotic dynamics
occur. These populations are
represented on the Main Wound Control Loop.
The summation-comparator-detector node, the inflammation error signal,
and the macrophage controllers represent the acute inflammation population. The mesenchymal controlled load and their
output represent the wound module population. Why does the CAP wound get into a state of unhealed dynamical
stability, even one that is counter-productive to health? When the control loop runs without
perturbation or retriggers, when its populations interact without abnormal
dependencies such as predation or deprivation, then the loop runs clockwise
with smooth Verhulst style one-shot dynamics.
Under those healthy circumstances, it runs toward one attractor, the
convergent healed wound. When various
perturbations or sustainers challenge the system, then it runs
counterclockwise as well, forward and backward at various times and segments,
with abnormal interconnects, bypasses, short circuits, and feedbacks that
sustain the operations of the loop. It
cannot enter the stable convergent attractor of normal healing. If primary disease or injury is
sufficiently active or strong, then it might enter the divergent attractor of
active ulceration. That state is
clinically undesirable, but it is an attractor, a locus of
thermodynamic (and therefore dynamical) stability. If it cannot enter the convergent
attractor, and if it is not on the divergent attractor, then the system can
settle on its third attractor, a state of sustained non-convergent
chaos. As an attractor, this too is a
dynamically stable place for the system to be. (The system, the wound, can enter phases of
higher free energy and intermediate status between attractors. These are the transitions from one state to
another, the induction of active disease, or the induction of wound healing,
transitions which occur when new energy or information is put into the
system, such as an infection, a flare-up of primary autoimmune inflammation,
a vascular thrombosis that restricts circulation, versus debridement of the
wound or treatment with anti-immune drugs or procedural revascularization.) It is under the conditions of sustained disease or injury that
abnormal population dependencies develop.
As presented in Part 2, sustained primary disease, inflammation,
immunity, thrombosis, allergy, etc. can all lead to abnormal population
admixture in both time and space. Once
acute inflammation and wound module get a prolonged look and handshake with
each other, then abnormal dependencies develop which can keep the control
loop running in abnormal directions or cycles. However, the whole system is robust enough
that if primary disease and injury are fully abated and wound conditions
restored to reasonable anatomy and timewise or spatial organization, then
Verhulst sigmoidal one-shot dynamics can be restored, and the system can
again enter its convergent attractor.
This is what we are doing with our basic wound therapies, to keep
primary disease and injury under control, and to keep the main two
populations separated, thereby allowing the loop to run smoothly in a
clockwise direction, which is often sufficient to allow the wound to heal. The problem gets even worse when a third population appears,
chronic inflammation. This takes the
place of sustained primary injury, and becomes the key element that
perpetuates the intrinsic or self-sustaining chaotic dynamics of the loop. More on this on the next panel. In the first paragraph of the previous slide, we talked about “
. . . components ramping up then decaying as they complete their appointed
tasks . . . “ This was a bit erroneous
because the cells and populations do not really have appointed tasks. Those are teleological attributes that we
ascribe to them. The reality is that they
just function according to the basic principles of cell biology – genomics,
proteomics, ligand-receptor interactions, cytokines and signaling, mobility
and secretory responses, etc. They
just do their own thing – they just function.
The aggregate correct responses occur, and the correct structures
form, because they are self-organizing automata. Their few rules of interaction allowed to
operate without interference result in parsimoniously correct
morphology. That is the sole purpose
of the wound module repair population, to organize into a specific anatomical
structure. When normal dynamics are
upset or obstructed, then that self-organization cannot be achieved, and the
final intended structure, a reorganized stroma and healed wound, fails to
form. That is how the control loop
“knows” what the system reference is.
It doesn’t need to know. As the
correct structures form, passively-parsimoniously-automatically, then
variances from normal get smaller, and the reactive response of inflammation
is mitigated because there is less stimulus to turn inflammation on. The system reference is just a well of thermodynamic
stability and minimum free energy that occurs as the system elements, the
“cellular” automata of self-organization, find their basal state of assembly. One of the crucial issues to keep in mind about the failing
non-healing wound is that the cells and cell populations are normal, acting
and reacting as expected according to their genetic program. In the chronic chaotic non-healing wound,
whether extrinsically (2-population) or intrinsically (3-population)
pathological, not one of the cells is itself intrinsically pathological. Remember, these are not divergent wounds -
not ulcerating and getting worse. They
just are not healing. What is
disrupted is only their aggregate organization and self-assembly. In fact, in comparison to other cells and
organs, wound cells and populations have remarkably few intrinsic diseases or
genetic defects. They remain
inherently healthy and functionally correct through thick-and-thin, “failing”
only when they are responding normally and correctly to disordered inputs or
environment. This point is discussed
further in the addendum after the next slide. The wound is a system and its aggregate behavior must be studied
that way. However, the individual
elements in the system are tangible physical and chemical and biological
items which must obey all rules and principles of physics, chemistry, and
biology. The interactions,
dependencies, failures, and assembly of these various elements are based on
specific genes and chemicals and cell responses, the stuff that conventional
bioscience research has been able to elucidate and characterize for
bazillions of individual cell structures and biochemicals. These are the nuts and bolts, the wires and
components, glue and nails, the grease and gasoline that keep the wound
healing machinery moving and building. System physics and non-linear dynamics
explain how they interact in time, and how their aggregate behavior succeeds
or fails, but it is still necessary to understand how these broad events are
implemented or actuated by tangible physical components. These nuts-and-bolts items and interactions
must equally be known for an understanding of wound failure to be
complete. How it is that specific
wound healing cells, chemicals, and interactions are degraded, disorganized, deprived,
& denied are a matter for conventional biosciences to continue
discovering. This brings us back to our primary points. (1) When the wound, host,
and system are intrinsically healthy, then recurrent primary injury &
thrombosis are needed to perpetuate the wound control loop. (2) When intrinsic chronicity occurs, the wound
system is locked into a persistent self-sustaining chaotic attractor without
needing new inputs . . . |
|
|
|
|
|
This series of papers is meant to elucidate what the intrinsic
disorders of wound healing are. As
presented in the addendum below, wound healing cells, angiocytes and
fibroblasts, are robust, well-tested, and largely error free, with extremely
few genetic errors or primary diseases.
When the wound system goes wrong, it is because these healthy cells
get confused about how to interact and assemble because they are getting
barraged by abnormal inputs. The
abnormal inputs may be extrinsic to the core dynamics of the wound, or they
may come from within. Acute wounds. First,
consider the normal healthy wound.
This is due to incidental or one-time injury – a cut or scrape, a
burn, a surgical incision, a myocardial infarction. The entire wound response and repair system
is then triggered into existence. If
there is no further injury, then there is a smooth uprise and then decay in
the logistics and dynamics of the response.
The uprise in the acute inflammation and repair populations, from zero
to peak, follows sigmoidal (linear) Verhulst dynamics. These populations then decay as their
controlled tasks complete. The
coordinated behavior of these populations is a series of sequential
one-shots. The result is a
parsimoniously reassembled stroma with epithelial sequestration, i.e. normal
wound healing. Problems happen when
something sustains the system, some set of repetitive or persistent injuries
or disease that keep re-initiating or amplifying these processes. Chronic wounds – extrinsic (2-population chaos). Wound chronicity begins with
persistence of disease or injury. This
could be the prolonged action of untreated disease (inflammatory, thrombotic,
immune, allergy, traumatic, etc), or it could be disease or injury that
intermittently relapses in spite of treatment. This even includes a wound that may no
longer be subject to the original event, but which is in a sustained state of
acute inflammation due to neglected or improper care, i.e. the “uncontrolled
wound”. The dynamics of extrinsic
chronicity reflect several possible states.
First, the wound may not be chaotic, but rather divergent, i.e. actively
sick and ulcerating due to the effects of sustained injury or disease. Alternately, it may be on the convergent
attractor, actually healing, but doing so slowly with only minute changes
during short intervals of observation.
These scenarios are akin to those regions on the Mandelbrot set where
the system is near the chaotic boundary, and thus takes a long time to arrive
at the definitive states of divergence or convergence. The key point though is that extrinsic
chronicity reflects a state of 2-population chaos. As we know from basic principles of
non-linear population logistics, even two contingent populations can have
chaotic dynamics, sustaining and promoting each other while concurrently
suppressing, depriving, or predating each other. The two populations here are acute
inflammation and wound module. In the
normal one-shot acute wound, these two populations stay separated by time and
space. Their admixture and abnormal dependencies
develop only as a consequence of sustained primary disease or injury. When the wound, host, and repair system are intrinsically
healthy, then recurrent primary injury-disease-thrombosis-etc are needed to
perpetuate the wound control loop. The
inverse is true too, that if primary stressors or promoters are abated, then
the control loop is not perpetuated and it can settle. This means that clinically, CAP wounds due
to extrinsic correctible causes are generally easy to heal. They are suppressed or subverted by acute
inflammation, but acute inflammation abates after primary disease and injury
and other sustained stressors are relieved.
This of course is what happens in wound practice day in and day
out. Basic hygiene and topical care
are initiated, inflammation and edema are relieved, then causative diseases
and risks are corrected. Once these
basic milestones are achieved, the inhibitory population of acute
inflammation can no longer suppress or attack the repair population. Once relieved, wound dynamics settle back
to one-shot profiles, chaos disappears, and normal repair dynamics pick up
again. The problematic
difficult-to-heal wound occur when intrinsic 3-population chronicity ensues. Chronic wounds – intrinsic
(3-population chaos). Intrinsic
chronicity occurs in association with the appearance of a third population – chronic
inflammation. With extrinsic
chronicity, outside forces need to be active to sustain abnormal
behavior. When intrinsic wound chronicity
occurs, the system gets locked into a persistent attractor without needing
new inputs. Primary external diseases
and injury can be removed, yet chaotic and sometimes even divergent dynamics
will persist. Once the wound becomes
autonomous and self-perpetuating, it is no longer easy to heal, because the
inhibitions and negative dependencies of the third population cannot be so
easily removed or reverted. As all
wound clinicians know, these wounds are in fact exasperatingly difficult to
heal. As for the extrinsic wound, problems begin with
sustained primary disease or injury. Underlying
primary inducing disorders are those of chronic inflammation (reactive &
induced), infection-immunity, trauma-injury, allergy-atopy, and thrombosis
& micro-occlusion.
With prolonged admixture of acute inflammatory cells, transformed
inflammatory cells (macrophage-histiocytes), early incidental lymphoid cells,
and repair cells and products, the
risk is that sooner or later auto-immunization will occur against repair
cells (angiocytes and fibroblasts).
The third population of chronic inflammation appears. Since it is explicitly tied to the repair
population, these two populations cannot be so easily separated, making the
third population intrinsic within the wound.
This third population brings with it predatory dependencies and
suppressions which subvert the functions of the repair population. Without specific intervention or treatment
to correct this situation, that’s that, and repair is fundamentally put out
of commission. It is quite interesting to read extant literature on the subject
of peri-vascular lymphoid infiltrates.
It is seen with all of the pathological associations that we might
expect after taking a look at this subject from a wound and immunopathy point
of view. Nonetheless it is not
recognized for what it is, nor its association with wounds and impaired
healing, even though the nature of lymphoid cells are well understood. Here are a few quotes / paraphrases from
relatively recent journal articles: “ women 20-34
years - infiltration of vessel wall by mononuclear cells, mainly lymphocytes
- 4 patients antiphospholipid antibodies - one heterozygous factor V Leiden
- we term this lymphocytic
thrombophilic arteritis “ Lee JS,
Kossard S, McGrath MA. Arch Dermatol.
2008;144(9):1175-1182. “ criteria for
lymphocytic vasculitis (1) lymphocytic infiltrate blood vessel, (2) fibrinoid
necrosis, (3) endothelial cell hyperplasia - clinical diagnoses varied - drug
reaction, chronic urticaria, nodular scabies, erythema multiforme, - probably not specific - more likely a
reactive process “ Massa
MC. Journal of Cutaneous Pathology,
2006, 11(2): 132–139. “ lymphocytic
vasculitis as sign of early connective tissue disease - hemorrhagic acral
livedo with anti-ribonucleo-protein antibodies - striking lymphocytic
vascular reaction - , perivascular extravasation of red cells,
leukocytoclastic debris - finally developed systemic lupus erythematosus “ Oh CW,
Lee SH, Heo EP. Am J Dermatopathol.
2003 Oct;25(5):423-7. “ lymphocytic
vasculitis - correlates with broad clinical differential diagnosis -
connective tissue disease (mostly sle), rickettsia and viruses, idiopathic
lichenoid dermatoses, perniosis, ulcerative necrotic mucha-habermann “ Carlson
JA, Chen KR. Am J Dermatopathol. 2007
Feb;29(1):32-43. Pathology textbooks have varying amounts of information on the
subject of lymphocytic vasculitis. One
of the more thorough discussions is in the large dermatopathology textbook Skin
Pathology (Weedon D. 2nd edition 2002 Churchill Livingstone.
pp 242-253, and nearby chapters). In
all of these writings, the emphasis is on describing histological features
and correlating them with eponymic clinical syndromes, a “dead poet’s
society” of dermatology. There never
seems to be an appreciation of underlying core pathophysiological principles
and mechanisms. The problem is that
wound healing is not recognized as a part of pathology, and wound pathologies
have little or no recognition or appreciation among pathologists,
dermatologists, and others who have written papers such as above (see Part 2,
slide 59). One of the common mistakes
behind this failed appreciation is that technical and vernacular meanings of
the word “chronic” are often confused or ignored. Histologically, chronic inflammation
appears as granular leukocytes (eosinophils, especially with allergic and
atopic conditions) and as lymphoid cells (lymphocytes and plasma cells). There is a tendency for pathologists to
issue wound reports that just say “inflammation” or “acute and chronic
inflammation” or something along those lines, with no further comment (your
experiences with your own colleagues in Pathology may vary . . . but I doubt
it). Comments like this trivialize the
identity and functions of the chronic inflammatory cell set. They mislead people into seeing chronic
inflammation as just a long standing substitute for acute inflammation. “When inflammation starts, it is early, and
therefore ipso facto it is acute . . . but when it hangs around long enough,
then we can call it chronic because it has been there a long time.” See how technical and vernacular definitions
are getting confused in that kind of implicit thinking? (Perhaps the term “chronic” should be
ditched from the lexicon of inflammation, using instead “lymphoid
inflammation” or “lymphoid infiltration” to eliminate any confusion or
misinterpretation.) The reality is
that chronic inflammation is entirely different than acute inflammation in
terms of biological functions and implications. All cells have specific parsimoniously defined biological
functions. This includes “chronic
inflammation” lymphoid cells, the 3rd population of the pathological wound. In other words, all cells have explicit
functions. The reason that there is so
much differentiation or “speciation” among the various cells, tissues, and
organs of the body is that one cell can only do so much. Each cell has a major function, such as
secreting mucus or making thyroid hormone or contracting when triggered by a
nerve. Granted, many cells have a
number of ancillary or collateral functions, but the differentiation and
functions of a cell are still rather narrowly and efficiently defined. Lymphoid cells are the agents of
immunity. They do not show up in the
wound just because, not on a lark, not on a whim, not as a matter of
confusion, not for a vacation nor a change of scenery, not in the rain and
not on a train, and not in a box and not with a fox (with apologies to
Theodor Geisel, Dr. Seuss). And, by
all means, they are NOT the relief team.
They do not get sent into the game because the first team is getting
tired and needs a break. This isn’t a
case of extra innings. Lymphoid cells
have an immune function. If they show
up in a wound, it is because some type of immune-related taxis, tropism, or
induction is attracting, guiding, or maturing them there. They are attracted to angioid and fibrous
elements of the stroma – that is what they hug once they are in the
wound. This should not be surprising because
that is what they get exposed to during prolonged admixed acute inflammation
and auto-sensitization. Once they have
this automatic attraction to stromal cells and structures, they are now part
of the stroma and the wound. Their
effects have become intrinsic in the wound, and they are presumably inhibitory effects. In normal healthy acute wound healing, each cell – monocyte, angiocyte,
fibroblast, keratinocyte – has an “assigned” job, a parsimoniously defined
task. If they can all just do their
jobs, then the wound self-organizes back to a stable stroma. When you look at a chronic and pathological
wound, productive self-organization is not happening. However, from the point of view of a single
plasma cell, monocyte, or fibroblast, there may be nothing really wrong, just
“life in the ‘hood”. As automata, they
have no insight or collective concept of what they are trying to build. Instead, as long as they are alive, they
just do their own thing, day-by-day, reacting as programmed to local
stimuli. And as long as local cells
can indeed do their own thing, then the wound and stroma reorganize without
problem. Problems happen when you
throw a cadre of chronic inflammatory cells into the neighborhood. Are they good cops, bad cops, street thugs
and bullies, misguided vigilantes, officially sanctioned law enforcement,
civil defense, or sanitation workers to clean up the mess? It depends on your point of view, but if
you are a neighborhood angiocyte, then that lymphocyte who doesn’t live on
your street is probably going to beat you up.
Although each cell is alive and functioning correctly, reacting
properly to local events, the collective system and its set of cells fails to
organize. In acute and in extrinsic 2-population wounds, primary disease
and injury sustain the wound. When
acute inflammation can be eliminated, wound healing returns to normal. Once the third population and its dynamical
dependencies and attractors have settled in, then they themselves sustain pathology. The third population is just another form
of challenge or injury which can perpetuate problems. The difference though is that the third
population is attracted to the repair population, and in so being it is
intrinsic in the wound and cannot be eliminated by treating or relieving
primary disease. Because of the third
population’s intrinsic status within the wound, this becomes an inherently
stable dynamical state that is hard to revert. |
|
|
|
|
|
Addendum to
Slide 34 Above, we mentioned that wound healing cells are robust,
well-tested, and largely error free, with extremely few genetic errors or
primary diseases. There are
extraordinarily few native diseases of the stroma. Those that do occur are largely autoimmune
in nature. We have seen in Part 2 why the
connective tissue disorders are due to autoimmune states as opposed to some
other general class of pathology. We
have also established the connection between autoimmunity and the diseases of
the general stroma. However, we have
yet to answer the other side of that question: why are there no common
diseases of the fascias, connective tissues, and general stroma related to
metabolic alterations or genetic deficiencies? The full annotated answer is beyond the
scope of this presentation, but a few basics can be explained. The stromal cells, fibroblasts and angiocytes, represent
evolutionary and phylogenetically ancient cells. Multicellular life appeared about 1 billion
years ago, as single celled life learned that there is strength and survival
advantage in cooperative association and the division and specialization of
labor. There are two quintessential
constructs needed to permit multicellular association and function: (1) some
system for holding everything together in a stable functional anatomical
form, and (2) some sort of distribution system to permit the interchange of
nutrients, metabolites, and information.
In animals, the system that evolved for holding things together is
based on connective proteins, the most abundant of which is collagen. Collagen structures and anatomy became
increasingly complex as life advanced, but collagen is present even in the
most primitive of multicellular organisms, the Porifera, the sponges. Evidence of a bulk transport system – a
vascular system – is also seen in some sponges, and it is permanently
established by the Cnidaria, the hydras and jellyfish. Primitive invertebrates do not have a blood
circulatory system. Instead, their gut
has extensions into all parts of the organism to directly deliver food, a
gastrovascular cavity that handles both digestion and distribution. Nonetheless, this is a vascular
distribution network, and our blood circulatory vascular system is a direct
evolutionary descendant of the gastrovascular cavities of the Cnidaria. Only one gene and its product are required
to govern the formation and morphology of this vascular distribution system,
and that gene is VEGF (vascular endothelial growth factor; well, actually 2
genes, VEGF and VEGFR, its receptor).
Genetic sequencing allows us to recognize the specific nucleotide
“spelling” of each gene, and jellyfish and human VEGF and VEGFR are highly
homologous, spelled almost exactly the same.
Also, the observable functions of VEGF on vascular cells and
structures are identical for jellyfish and humans. As life evolved, many new genes appeared,
old ones disappeared, and many morphed and changed. But, over eons of multicellular evolution,
VEGF and its functions are unchanged.
Why? Why has VEGF remained unchanged?
Because multicellular life is wholly contingent on a bulk transport
vascular distribution system. Without
it, complex multicellular life is categorically impossible. (Likewise for the proteinaceous support
matrix that structurally binds organism together.) Once this core infrastructure element of
life had been written, it needed no revision, because it worked so well. What this means is that for the few
quintessential genes that permit multicellular life, there is little room for
mutation. VEGF is so crucially
essential for life that without it, an embryo unconditionally cannot develop
– period - exclamation. (In some
experiments, VEGF knockout is categorically lethal; in other experiments,
other angiogenic factors can keep a conceptus alive, but with significant
developmental defects.) Whatever VEGF
mutation might occur in a gamete, it cannot be propagated, because a
conceptus simply cannot develop beyond just a few cells (the gastrula
stage). The basic stromal structure of
multicellular life – connective matrix and vascular distribution system – was
worked out from the beginning, 1 billion years ago. The formation and function of these
structures and cells has been thoroughly tested and debugged, meaning they
are essentially error free. These core
infrastructure functions of multicellular life are so consistently conserved
and dependable, so thoroughly robust, that there are no major genetic or
metabolic disorders of the stroma, and consequently none of the mesenchymal
component of wound healing. Because
these cells and structures have extraordinarily few intrinsic disorders, when
wound healing goes bad it reflects some sort of exogenous disorder or damage,
some sort of deprivation or attack affecting these cells and structures. That includes non-specific non-targeted
conditions such as trauma, ischemia, toxicities, and severe
metabolic-nutritional inadequacy. It
also includes targeted damage directed against these cells and structures,
and as we have seen in this section, that means the auto-immune disorders. |
|
|
|
|
|
These diagrams summarize the points of the last slide. On top is the Main Wound Control
Loop. Some of the blocks have been
corralled into two major groups, representing the two essential populations
of wound healing, acute inflammation and wound module. In addition, exogenous injury is summed
into the loop, as explained in Part 1.
Obviously, ongoing primary injury will keep re-elevating the state of
the wound, even as the control loop is otherwise trying to wind it down. This is the basis for 2-population
extrinsic chronicity and chaos (which can also eventually spawn the third
population). Below right is the graph from slide 26 showing the dynamics
that occur when chronic inflammation is added. Recall that in Part 1 it was stressed that
the control loop is an open model that can represent all states of wound
physiology and pathology. Below
left, to keep consistent with the control model, chronic inflammation and
the smaller graph are melded into the main loop. The chronic inflammation population has
inhibitory effects on the wound module population, and promotional effects on
the acute inflammation population, as well as promotional effects on the
state of the ulcer. If ongoing acute
injury or disease were also added, then that would be another layer of
control, making the dynamics yet more chaotic. Notice that chronic inflammation occupies a
similar position as active injury, and it serves the same dynamics of
sustaining ulceration, acute inflammation, and the ongoing operations of the
loop. The difference is that extrinsic
disease and injury can almost always be abated, allowing the loop to settle
into the desirable state of one-shot dynamics, loop activity decaying as the
wound converges on its fulfilled state of closure. However, chronic auto-immunized
inflammation becomes intrinsic within the wound or stroma, and it cannot be
so easily abated. As such, it acts
like a generator or pacer to sustain wound injury and acute inflammation, to
suppress repair, and in so doing perpetuate ongoing chaotic non-convergent
wound dynamics. |
|
|
|
|
|
We have seen that there are several primary inducing disorders
that can perpetuate acute inflammation (and thus the wound control loop),
increase auto-antigenic load, and risk auto-immunization by lymphoid
sensitization. These are all disorders
of chronic, recurrent, persistent, sustained injury, inflammation, and
thrombosis, including primary inflammation (reactive & induced),
infection-immunity, trauma-injury, allergy-atopy, and thrombosis &
micro-occlusion. This slide
illustrates the effects of primary disease or injury to perpetuate acute
inflammation and potentially induce a state of auto-immunized chronic
inflammation. Left: a leg ulcer from a
patient with polyarteritis nodosa.
This view is from below an area of acute active skin infarction and
ulceration. Vessels throughout the
area were infiltrated by acute neutrophilic inflammation. It is this state of persistent or
repetitive acute inflammation which chews up cells and releases endocellular
debris and antigens. While this
particular specimen had few chronic inflammatory cells, this would seem to be
the type of patient prone to stromal auto-immunization, and quite likely it
occurred long ago and is the basis for repetitive acute inflammation. Right: This patient developed acute progressive ulceration
of the ankle after biopsy of a small lesion.
She had protein C deficiency and positive cryoglobulins, along with
histologic findings of thrombosis and vascular necrosis. In a patient with a primary hypercoagulable
disorder, it is sustained small vessel thrombosis which can create ongoing
low level acute inflammation as well as cause angiocyte necrosis and the release
of related cellular debris. |
|
|
|
|
|
This is another example of the effects of unrelenting primary
disease to induce sustained acute inflammation. These images are from an 31 year old woman
with a primary hypercoagulable disorder and ankle ulcer, along with
miscellaneous immunopathic symptoms.
Laboratory evaluation confirmed low protein C and low APC resistance
(likely the primary problems), a positive lupus anticoagulant (probably
secondary due to induced auto-immunity) and low factor IX (indirect evidence
of a thrombotic state, due to compensatory down-regulation of a prothrombotic
element). The specimens are from the
base and edge of the ulcer. Features
that can be observed include: acute
and chronic inflammation, acute and chronic thrombosis, vascular necrosis or
fibrinoid degeneration, wound surface infarcts, vascular disruption and
disorganization, dense peri-vascular plasma cell and lymphocyte infiltration,
cellular debris and basophilia deeper than expected for healthy wounds,
cellular debris and basophilia along angiogenic cords, scant or disorganized
fibroplasia, and an admixture of acute inflammation. wound module, and
chronic inflammatory cells at different levels or strata. As seen in the gross picture, there is active inflammation and necrosis
in spite of treatment, representing
persistence of the pathological state.
Histologically, the overall architecture of the wound module is
relatively correct, but numerous features are altered from normality. In this case, the primary hypercoagulable
state is responsible for the persistent state of active infarctive and
inflammatory pathology, which will in turn inhibit and delay wound healing,
as well as create the conditions for auto-immunization. Recognizing an auto-immune component of
delayed or disrupted healing becomes easier only after non-immune acute
events have been controlled. However,
in this case, the active pathological state was not controlled by
anticoagulants and topical care alone, and persistent auto-immune
inflammation may have been the persistent promoter of continued
micro-thrombosis. Untangling the
interconnections of acute inflammation, chronic inflammation, thrombosis, and
their effects on subsequent wound healing
becomes difficult, because this whole interconnected mess IS the intrinsic
disease of wound healing. |
|
|
|
|
|
This and the next few slides show examples of the third
population, chronic inflammation, in wounds due to one of the typical chronic
inducing disorders. These images are
from two patients with primary hypercoagulable disorders. These views near the edges of the ulcers both
show a vascular locus, i.e. a zone of vessels and angioid tissue within a
dermal or connective matrix. The vessels
have chronic thrombi and reorganization.
The specimen on the left shows migratory spindle shaped
disorganization of the angiocytes consistent with a state of chronic
inflammation and nominal “wound healing”, i.e. growth factor induced
mesenchymal cell activities. In both
specimens, the vascular locus is heavily infiltrated with chronic
inflammatory cells (lymphocytes, plasma cells, eosinophils). The left specimen is infiltrated with
almost pure lymphocytes. The right
specimen is infiltrated with mostly all plasma cells and eosinophils. There are few neutrophils, and the
surrounding collagen matrix shows normal fibroblasts without inflammatory
changes (that would have been much different if the specimens came from
directly within the ulcer). Why are
these vessels in a state of chronic immunogenic inflammation? |
|
|
|
|
|
These images are from a 55 year old woman with chronic ulcers of
the abdomen. She had a long history of
chronic recurrent ventral hernia with numerous failed operations, and chronic
abscess around polymer mesh. The mesh
was ultimately removed, and the remaining open wound eventually healed by
progressive contraction and epithelialization. As the patient neared the end of complete
closure, she suddenly developed inflammatory changes and ulceration in areas
of regenerated epithelium that had been healed and seemingly healthy for many
months. Some of these new ulcers
stabilized and became persistent and non-healing, whereas others continued
slow progressive enlargement.
Histology shows a rich mix of acute inflammation, chronic inflammation
and wound repair. Left: this specimen is at the margin of
ulceration (epidermal edge just a few microns away to the left). There is angioid proliferation consistent
with wound healing events, but the vascular locus is completely filled with
lymphoid cells. The epidermis above is
undergoing active lysis by neutrophil invasion. The assumption is that the lymphoid cells
have triggered a new round of acute lytic inflammation. This is a paradigm of the
inflammatory-lytic pattern of necrosis and ulceration. Right: migratory spindle-shaped angioid cells
would at face value appear to be normal, migrating chemotropically in a
gradient field of angiogenic factors near the surface of the wound. Yet they have attracted the attention of a
bevy of plasma cells. Within that intermix
zone, the normal vertical migratory pattern of the angiocytes has been
disorganized. |
|
|
|
|
|
These images are from a 53 year old woman with lupus since age
18. She had had multiple abdominal
ventral hernias and failed repairs.
The current specimen was from a recent incision or scar that
spontaneously started to erode and re-ulcerate. Left: at the edge of the ulcer, in the zone of
active lytic ulceration, is a coagulum that is overrun with an equal mix of
acute inflammation, nuclear debris, lymphoid inflammation (plasma cells and
lymphocytes), and repair cells. This
lesion is actively ulcerating, so repair cell proliferation is futile,
non-productive, non-constructive, but their presence indicates that these
cells are all attempting to do their nominal job. Regardless of the histologic presence of
angioid cells, the wound was actively ulcerating (divergent), far from being
able to organize and heal. Right
top: a reorganizing set of vessels
in the vertical migration zone in the upper strata of the wound. Here too is a mix of chronic and acute
inflammation and wound module proliferation.
The chronic inflammatory cells have an affinity for the angioid
cells. Right bottom: a vascular cluster deeper in the dermis, in
which lympho-plasmacytic infiltration is very dense. This was typical of most of the vessels in
the specimen. |
|
|
|
|
|
The left and center images are from a 34 year old man who was
quadriplegic following spine injury. He
presented with literally thousands of individual ulcers on trunk and
extremities ranging in size from 1-2 mm through many centimeters. The active phase of ulceration began with
eosinophil rich skin bullae, so the working diagnosis is bullous
pemphigoid. No lesions would heal with
generic topical therapies. Healing began
with systemic steroids, but was only partial.
Rapid complete healing was induced with azathioprine, which more or
less proves that the chronic inflammatory population was the culprit behind
the wound failures. Left: a vertical view of the wound showing a
largely normal gross architecture, but with excessive edema and/or
aminoglycan. Near the bottom are dark
basophilic infiltrates which are almost all plasma cells. Clinically, the wounds were not
healing. Histologically, although
repair cells are present, there is no fibroplasia and incomplete organization
of the stromal elements. Center
top: a non-specific area showing
intense admixture of acute inflammation, nuclear debris,
histiocytes-phagocytes, lymphocytes and plasmacytes, and angioid reparative
cells. Center bottom: Acute and lymphoid inflammatory cells
surrounding and infiltrated into a vessel.
The vessel itself is highly disorganized, the angioid cells only
marginally associated or interconnected, and obviously leaky as evidenced by
the significant hemorrhage throughout the area. Right top, bottom: These images are from refractory leg ulcers in a 53 year old man
with rheumatoid. The tissues are dense
with plasma cells as well as acute inflammation, intermixed with the
angiocytes and fibroblasts of the developing (or not developing) stroma. Although
the repair cells are present, they are failing to fully coalesce and
stabilize. There is stasis and
leukocyte trapping within the vessels.
All three dynamical populations are here, acute inflammation, chronic
inflammation, and wound module. They
are admixed at multiple strata of the wound, and none of them appear as they
should in a healthy wound. |
|
|
|
|
|
These four images are from chronic and non-healing wounds in 4
different patients with various chronic antecedent injury or disease. They all show an admixture of acute
inflammation and repair cells with chronic lymphocytic inflammation. Left top: from a chronic sacral pressure ulcer. Pressure ulcers are due to a trauma. The cause of the wound is strictly
extrinsic to the wound healing process, and such wounds are expected to have
normal wound healing kinetics as long as pressure and other adverse mechanics
and contact conditions are relieved. This
is generally true, but there is also nothing unusual about a chronic pressure
ulcer that seems to defy these expectations, acting impaired and difficult to
heal even after topical conditions are completely relieved. These patients obviously have globally
normal wound healing, and you can do surgery elsewhere on their bodies
without problems, but the primary wounds seem to be misbehaved. The key element in this adverse
transformation to an impaired wound may just be the state of chronicity
itself, giving the wound a chance to develop some degree of
lymphocyte-mediated auto-sensitization and disruption of the dynamical
integrity of the wound module. This
specimen is from the angio-attraction and angio-organization aminoglycan
strata of the wound, and it would appear fairly normal except for one thing –
the intense plasma cell infiltration along the vascular locus and elsewhere
where angiocytes are streaming. Left bottom: from a
prolonged (years) ankle ulcer following radiation for a melanoma. With various treatments, this wound several
times was nearly healed, but then re-ulcerated with no apparent
provocation. Virtually every vessel in
her specimens, whether under the ulcer or beyond its limits, had an intense
wide lymphocyte infiltrate. On the
wound surface (pictured), lymphocyte and acute inflammation are strongly
mixed. Lymphocytes are especially
intense around angioid structures (which elsewhere in the specimen are
somewhat dysmorphic and disorganized). Right top: from a
patient with rheumatoid arthritis.
This is another good example of the admixture of all three populations
below the surface of a refractory chronic ulcer. Plasmacytes and lymphocytes are clustered
in and around vessels and angiocytes. Right bottom: from a 35
year old woman with a years-long refractory leg ulcer of uncertain primary
diagnosis. Histology shows features of
many CAP and immunopathic wounds:
neutrophilic peri-arteritis (in a grossly bland uninflamed wound),
plasma cell and eosinophil infiltrates, vascular disorganization, narrow or
thin zones of aminoglycans and angio-organization, insignificant or
disorganized fibroplasia, dense nuclear debris. In this specimen, the population admixture
was intense, but also intensely disorganized.
There were swarms of angioid cells, but not a lot of well-formed angiogenesis
or coalesced vascular structures. In
addition, there were an extraordinary number of angioid mitoses, present in
most high power fields, sometimes 2 or 3, almost what you could expect to see
with anaplastic cancers. These angioid
cells were proliferating as they would in any healthy wound, but with
bizarrely excessive turnover. Yet grossly
the wound had only the scant appearance of “granulation tissue”, and it was
largely absent histologically (failed organizational angiogenesis). One of those mitoses is seen here, in the
midst of all the mayhem. If there was
so much angioid turnover, yet no significant angiogenesis, what was
happening? In cancers, new cells
appear and accumulate. Here, they would
seem to be rapidly generating then wholly disappearing. Possibly they were destroyed by acute inflammatory
cells or else by immune mediated events.
Perhaps they died by apoptosis, perhaps something else. Either way, they would leave behind a lot
of potentially antigenic debris that could further the antigenic recognition
and auto-sensitization against angioid cells and structures. The mitotic rate aside, the presence of
chronic inflammatory cells, peri-vasculitis, altered behaviors of angioid and
fibrous cells, and corruption of normal wound strata makes this a typical
pathological wound, almost certainly of auto-immunopathic origin and
intrinsic chronicity. |
|
|
|
|
|
Addendum to
Slide 42 We began Part 3 by asking what is the quintessential structure,
function, and derangement of the wound.
As we have seen, it is an ad hoc reserve organ made from a set of cell
populations doing interactive things for the sake of reorganizing its own
structure. Failure to organize is a
derangement of the non-linear inter-operations of its normally controlled
elements and constituent populations, i.e. a “dynamical disorder” or
“dysdynamia”. “Dynamical disorders” are getting attention from people who
study a variety of complex systems in biology, because they affect
cardiovascular functions, neurological functions, endocrine and nutritional
functions, even population and herd dynamics, and everything else. As we saw in Part 2, it is the autoimmune connective
tissue disorders (whether primary and a priori versus induced in the wound by
chronicity and cell population admixture) that are the diseases which affect
the intrinsic elements and individual components of the wound healing
system. Dysdynamia is the disorder
which affects their inter-operations and the collective function of the
system. In principle, recognizing the auto-immune or intrinsic
chronicity component of delayed or disrupted healing becomes easier or
diagnostically accurate only after injury, disease, and non-immune acute
inflammation have been controlled.
However, untangling the interconnections of thrombosis and acute
inflammation, immunity and chronic inflammation, and their effects on wound
healing becomes very difficult,
because this whole entanglement IS the disease of wound healing. When the system gets entangled to the point
that these core events in stromal biology cannot function properly and the
wound module cannot converge, that is the intrinsic
disease-disorder-pathology of the wound.
Intrinsic wound chronicity is simply a non-convergent attractor of a
chaotic entanglement of contingent cell populations. This attractor is clinically undesirable,
the wound being seen as a refractory illness, but dynamically it is a stable
state from which it cannot escape without deliberate treatment. Every therapeutic intervention that is
tried is an attempt to unentangle the populations and dynamics of this
multi-control system, trying to put “energy” in that can raise its orbit
enough to shift to a more favorable convergent attractor. Stromal autoimmunity, a targeted disorder of angiocytes and
fibroblasts, is an essential inducer
and maintainer of this intrinsic wound chronicity and dysdynamia. However, these are just two elements of a
complex inter-operative system. What
about all of the other things that might go wrong with the wound or its
components, do they not also get sick and contribute to wound failure or
wound incompetence? No, not much. In spite of the complexity of the wound, in
spite of the bazillion individual micro-structures and chemicals that participate
in the wound healing story, no, not much else goes intrinsically wrong with
the wound. The reasons that there are
so extremely few metabolic or genetic disorders of the stroma are for the
evolutionary reasons discussed in the addendum to slide 34. However, the system has an Achilles heel. When the wound module is left to itself, it very reliably
self-organizes back to a stable re-epithelialized stroma. If there are no major genetic or metabolic
disorders of the stroma, and consequently none of the mesenchymal component
of wound healing, then when wound healing goes bad it reflects some sort of
exogenous deprivation or attack. Under
normal healthy circumstances, these extrinsic challenges come from the open ambient
world. Normal wound anatomy is
structured to provide its own protection.
The plasma protein and inflammatory layer on the surface of the wound
(with its host of neutrophils and related things) becomes the ambient
interface, the protective barrier, a quite reasonable and effective
substitute for missing epithelium. This
top stratum provides perfect shelter for the repair process underneath. (See the Arimedica website for detailed
information on normal wound histo-anatomy, especially the file arimedica_integra
histogenesis_gottlieb-me_v2003.pdf).
So, in principle, since the wound module elements are intrinsically
error-free, and since they are protected from exogenous attack from above,
then wound healing should work perfectly, and it does, except . . . Its Achilles heel is its unsuspecting soft
underbelly. The only thing that can
and does goes wrong is the unexpected auto-attack, a job from the inside. This is what happens when auto-immunization
occurs and the defender system is directed against the repair system. This attack from a misdirected population
is what disrupts self-organization. |
|
|
|
|
|
Our focus has been on the auto-immunization of the stroma and
the intrinsic dysfunction of the wound.
Intrinsic dysfunction occurs when everything in the control loop is
functioning correctly according to its own program, but elements are not
cooperating or organizing due to population & dynamical chaos. It is important now to remind that not all
chronic and pathological wounds are intrinsically impaired. Many are chronic (vernacular sense – there
a long time), because primary disease or injury or other extrinsic causative
factors remain active. Here are examples where non-healing is due to identifiable
extrinsic pathologies that directly injure or inhibit wound elements or
disrupt control loop operations: an
open tendon with open tendon sheath; a
pressure ulcer with open inter-tarsal joints;
an ulcer that starts and stops healing with cycles of
hydroxyurea; a radiation wound; unnatural iatrogenic injury due to topical
chemicals; an arterial ulcer over the
tibia which healed promptly upon restoration of circulation and placement of
a regenerative matrix (cadaveric dermis) so that repair cells would have a
place to do their business. These wounds are chronic and pathological, and the control loop
and system dynamics remain active. The
principles of chaos and dysdynamia apply.
However, for extrinsically pathological wounds, the dysdynamia results
from the sustained activity of injury and acute inflammation. This is the 2-population chronic wound, and
they are generally easy to treat once primary disease and injury have been
corrected. It is the obligation of the clinician to make sure that
diagnosis is correct so that proper treatment can be planned. |
|
|
|
|
|
This slide is a reminder that this is a three part presentation
that looks at wound pathology from the point of view of its applicable
physics, elucidating the intrinsic dysfunctions of the wound as a result of
dysdynamia, especially when stromal auto-immunization has occurred due to
prolonged population admixture in a repetitively injured wound. Part 1 – The Wound as a System and a Controlled Machine Part 2 – Auto-Immunopathy and the Intrinsic Disease of Wound
Healing Part 3 – Chronicity and the Physics of
Wound Failure These presentations and supplementary
materials are all available at www.arimedica.com . |
|
|
|
|
|
SUMMARY Chronicity, chronic inflammation, & the physics of wound
failure All systems – physical, biological, natural, engineered – must
obey basic physical laws and relationships, e.g. thermodynamics, conservation,
Newton, Ohm, Maxwell, physical chemistry, mathematics. The wound is no different. While conventional and classically trained
bioscientists focus on biochemical constituents of cells and matrix and
wounds, this approach to study cannot elucidate the inter-operational functions
and failures of the wound. As a
complex system, systems physics – aka non-linear dynamics – is required to
understand the integrated timewise operations and failures of the wound. The wound module is a
special ad hoc reserve organ. It
appears in response to injury, then vanishes when the wound is healed. As for any other organ or tissue, it has a
core structure and function, a core mode of failure, and an applicable
physics to explain its functions and failures. What are the quintessential structures and functions of the
wound? It is a collection of mutually
interactive self-organizing cell populations.
It has no other function than to organize itself into a generic stroma
to support epithelium and other tissues. What is the quintessential derangement of intrinsic wound
pathology & chronicity? It is
dynamical disorder of logistical self-re-organization among these
populations. When it fails, it simply
fails to organize to its intended final form, to complete its task to become
something and then disappear. What are the fundamental physics relevant to the wound? Non-linear dynamics, control, chaos,
population logistics, cellular automata.
It is the science of populations, their interactions, control, and self-organization. When it fails, it is a dynamical disorder
of self-organizing populations. |
|
|
|
|
|
SUMMARY The Physics & Pathology of Wounds - 3 Chronicity and the Intrinsic Disease of Wound Healing There are few inborn errors of wound healing. What then is the intrinsic pathology of the wound? It is a state of autonomy or self-perpetuation not contingent on
the primary pathology. It is a dynamical disorder of a complex system due to: 1 – Continued primary disease, injury, inflammation, thrombosis which
leads to sustained acute inflammation.
This is a predicate condition which leads to stromal auto-immunization. 2 – Stromal auto-immunization leads to the appearance of
lymphocytic (aka “chronic”) inflammation.
This becomes the abnormal 3rd cell population in the wound. 3 – Sustained acute inflammation (the 1st population) and chronic
immune inflammation (the 3rd population) create population dependencies and
feedbacks that continue to sustain thrombosis and inflammation and dynamical
disorganization. They also create
conditions of predation and deprivation that inhibit or disorganize the 2nd
population, the repair module, thereby disrupting self-organization and
keeping the wound from healing. 4 – A complex multicontrol system such as this mix will have
certain dynamical behaviors or attractors: convergence (healing, the “healthy
wound”), divergence (active disease and ulceration, the “sick wound”), or
chaotic orbits (non-healing, the “impaired wound”). While the chaotic non-healing wound is
clinically undesirable, it represents a dynamically stable attractor and
thermodynamic basin that makes escape to a convergent attractor difficult to
achieve. Primary disease, injury, inflammation, thrombosis have a crucial
role in perpetuating the early wound and in inducing chronic inflammation. Underlying primary inducing disorders are
those of chronic inflammation (reactive & induced), infection-immunity,
trauma-injury, allergy-atopy, and thrombosis & micro-occlusion. Sustained primary disease and injury can lead to chaotic
dynamics in the 2-population wound, To
the extent that they continue, then the pathological non-healing state
remains active and non-trivial to break.
However, this state is generally easier to break than chaos in the
intrinsic 3-population wound. In the
extrinsic 2-population, if primary disease is controlled, which is almost
always possible, then the repair module can function without inhibition, and organized
wound healing resumes. Once the third population and its dynamical dependencies and
attractors have settled in, then primary pathology is no longer needed to
sustain the problem. The wound can
then persist even when good care has resolved acute stressors. Because the 3rd pathological population is
inherently linked to the others, the pathology and dysdynamia become
intrinsic, the 3-population wound.
These are the wounds that are neither healing nor getting worse. They go through perpetual cellular and
biological activity, but with no net gain.
The inherent dynamical stability of this state resists treatment. This inherent stability makes such wounds
clinically frustrating, but likewise they are safe and compatible with
otherwise healthy life. Stromal auto-immunopathy and 3-population chronicity are the
intrinsic diseases of wound healing.
The integrated timewise behavior of the intrinsically chronic wound
must be understood via the physics of complex systems and populations –
non-linear dynamics. The intrinsically
impaired wound is a dynamical disorganization of its mutually contingent cell
populations. Understanding the
intrinsically chronic wound via its physics correlates readily with clinical
behaviors and strategies for treatment. In summary, when you look at an intrinsically pathological
wound, chronic, difficult to heal, perpetually a bit better then a bit worse
in spite of treatment, you are seeing the standard dynamics of a complex
system as governed by the laws of the universe – physics. Dependencies between acute inflammation,
wound module, and chronic inflammation keep the wound module from fully
self-organizing. The linear dynamics
of a one-shot perturbation and response to a single trauma in a healthy
subject, i.e. normal wound healing does not exist in the chronic pathological
wound. Instead, the chronic pathological
wound exhibits typical non-linear dynamics, meaning chaos, orbits, and
clinically undesirable but dynamically stable attractors that keep the wound
module from getting to the finish line.
They go through perpetual cellular and biological activity with no net
gain. Wounds with intrinsic and
auto-immunopathic chronicity are simply behaving as complex natural systems
are expected to behave when their normal balance or attractor is stressed by
added populations, promoters, or inhibitors.
Such systems will tend to dwell in their attractors, for dynamical and
thermodynamical reasons, and breaking the abnormal population entanglements
and their chaotic orbits enough to get the wounds to heal requires deliberate
“strong” therapies. |
|
|
|
|
|
Epilogue FINAL THOUGHTS. The Physics and
Pathology of Wounds has been a triptych presentation meant to present a thesis
concerning the pathophysiology of the chronic and pathological wound. Several crucial points have been made
covering a diversity of subjects.
There are also a variety of loose ends and directions for further
study. In Part 1, The Wound as a System and a Controlled
Machine, the main point was that the wound is a system, that no amount of
conventional bioscience experimentation will elucidate the operational
physiology of the wound, that systems must be studied as such using the
relevant tools of physics and engineering,
We started by showing that the wound, like nearly all healthy
physiological systems, is a controlled system. Control makes sure that the wound healing
system stays asleep until needed, then comes to life, builds a
morphologically correct structure with just a few rules and cell types, then
winds down and reenters standby mode once its job is complete. All of the components of the wound response
and repair system have their place within the anatomical wound. They also have their place on a basic engineering
control loop that has feedback and a way to correct variances from a
reference state. The healthy wound and
all states of pathology can be modeled on this loop. Dynamical behaviors, healthy and altered,
can all be studied or understood from this perspective. In Part 2, Auto-Immunopathy and the Intrinsic Disease of
Wound Healing, we went from a physics and engineering perspective to a
clinical and pathological perspective.
“Rheumatoid” and other auto-immunopathic ulcers are a major class of
chronic and pathological wounds, and they are difficult to heal. CAP wounds due to other diagnoses can also
become refractory to care. The commonality
is that the wound is just connective stroma (angiocytes and fibroblasts,
vessels and connectives), wound healing is just the stroma reassembling
itself, and autoimmune connective tissue disorders are diseases directed
against the stroma. Whether patients
have a primary or an a priori CVD-CTD versus their wounds having become
auto-immunized and intrinsically chronic, it is just two similar pathways to
the same common state. For at least
some patients, such as those with primary hypercoagulable states, clinical
CVD-CTD’s such as rheumatoid or lupus are the consequence of stromal
auto-immunization from the sustained micro-thrombosis, making both the
CVD-CTD and the wound healing problems sequelae of the primary pathology. The intrinsically chronic wound is just a
phenotype or avatar or cultivar of the broad class of auto-immune connective
tissue disorders. The induction of
chronic lympho-plasmacytic infiltrates in the stroma correlates with
chronicity and intractability in wounds that will not heal, and it should
have a direct influence on treatments selected by the clinician. In Part 3, Chronicity and the Intrinsic Disease of Wound
Healing, the engineering aspects of the wound as a controlled process have
been combined with the clinico-pathological aspects of intrinsic auto-immune
wound chronicity to arrive at a physics based understanding of why the
disordered anatomy and cell mix of the chronic wound makes it impaired and
incompetent to heal. As a non-linear
dynamical system subject to feedback and control, it will exhibit chaos,
attractors, and n-body complexification.
Overall, its dynamics fall to one of three standard non-linear
dynamical states, convergence (healthy), divergence (sick), and orbiting or
chaotic (impaired). The healthy wound
has a separation of populations that ensures that each runs the control loop
as a self-converging one-shot.
Prolonged primary disease, injury, thrombosis, and inflammation create
conditions of progressive 2-population admixture (acute inflammation and wound
module) to the point that the immune system eventually “sees” and sensitizes
to stromal auto-antigens. The
subsequent induction and admixture of the “3rd population”, chronic
lympho-plasmacytic inflammation, then locks the system into a self-sustaining
chaotic orbit. Chronic inflammation
becomes a “generator” within the wound to perpetuate the acute state and keep
chaotic dynamics active, even after primary disease and injury have abated. These are the impaired intrinsically
pathological wounds, neither healing nor getting worse. They go through perpetual cellular and
biological activity with no net gain.
The inherent dynamical stability of this state resists treatment. Getting them healed requires persistent
treatment, with anti-inflammatory and anti-immune therapies topping the list
of crucial specific interventions. Part 4, A Gallery of Case Studies, was not given at the original
presentation of these papers, but is presented here to illustrate concepts,
patients and disease, and the direct utility of these concepts in making an
accurate wound diagnosis and picking proper effective treatment. If you are a classic bioscientist, it is likely that cell
biology, biochemistry, genomics, and the like are your “cup of tea”. Perhaps you are asking yourself questions
like “What are the specifics of how these cells interact with each
other? How do they signal each
other? How do they inhibit or promote
each other? Does one cell directly
suppress another, or does it ignore the cell and just block its excreted
output? What chemicals and genes are
on or off to implement these actions and reactions?” This third part of this series which explains
the physics of wound pathology and wound failure may seem novel or tangential
to these conventional “meat and potatoes” issues of biological research. However, the wound is a complex dynamical
system, and complex systems MUST be looked at this way to understand their
overall operations and failures. That
is one of the great accomplishments of science in the past 40 years, that
complex systems can in fact be studied and understood by non-linear dynamics,
making this the “century of the system”.
By focusing on the physics of the system, it is possible to see the
dynamics and operational inter-dependencies that govern how wound cells will
behave in the face of various normal and abnormal circumstances. In fact the physics of the pathological
wound simply do not care much about how these biological interactions are
effected, implemented, or actuated at a biochemical level. For example, growth factors are one of the
main elements in the control loop, but that is because they are a control
signal directly within the loop, However,
their exact chemical structure is completely irrelevant to their
system-integrated operational dynamics.
The physics do not care if an angiocyte has a certain surface antigen
nor if a fibroblast has a specific gene expression nor if an epithelial cell
has such-and-such intracellular signaling pathway turned on – no more than it
cares if the macrophages are wearing purple underpants. The physics of the wound is concerned with
system level interactions, control, non-linearity and chaos, population
interactions and logistics, thermodynamics and energy. It is equally true though that the tangible biology of this
system is equally important. It is
vital to understand the nuts-and-bolts, brick-and-mortar, collagen-and-matrix
alterations and failures that keep these populations from healing the wound –
what gene, what protein, what signaler, what receptor, what cofactor or
catalyst, etc. These are the issues of
classical bioscience that must be answered for the picture to be validated
and complete, and which must be answered in the biology laboratory. These are the discoveries or correlations
which are most likely to lead to the development of useful new treatments. What is needed now is bioscience research
to answer some of the concepts and questions presented here. The main questions to be investigated, based
on the novel observations and theses presented here, are how it is that peri-stromal
lymphocytes and plasmacytes exert their effect to retard or chaoticize the
wound. The nature and difficulties of “rheumatoid” wounds, their distinctive
patterns of inflammatory-lytic ulceration, their correlation with auto-immune
symptoms and endocellular auto-antibodies are all the stuff of everyday
clinical practice in dealing with chronic and pathological wounds. When I started this study, and started
looking very closely at the histo-pathology of these wounds, I had prior
assumptions about what I thought I would see.
I would see lymphocytes, plasma cells, and eosinophils, i.e. “chronic
inflammation” – I knew that already - that is just basic non-discriminating
casual freshman knowledge. What I also
thought I would see would be signs of aggressive immune inflammation –
antibody and complement fixation, killer T-cells, active lysis of angioid and
fibrous cells, with non-leukocytic mesenchymal debris all over the
place. Wrong. The lymphoid infiltrates are very clearly
associated with the active wound, always within fractions of a millimeter up
to a few millimeters from the wound surface or edges, but then tailing
out. Lymphoid aggregates my be a bit
deeper, but they are more isolated, more mature looking, and always isolated
to the vessels or vascular locus. And
that is another strong observation, that the lymphoid cells have a very clear
affinity for repair cells, especially the angioid cells and their vascular structures. However, they seem to coexist peacefully
enough. There are no signs of stromal
cell killing. I have done
immunohistochemistry on many of the samples seen here. There may be problems with the process and
specimen handling (it is not a research lab), so I cannot validate what has been
seen, but so far there has been little evidence of immunoglobulins or
complement, even when the slide is overrun with plasma cells. Nonetheless, these infiltrates are (1) associated
with wound chronicity and chaos, (2) they go away with treatment and then the
wounds heal, and (3) they are associated with a number of impairments and
disorganizational features in the wound histology. When it comes down to the more tangible biology and chemistry of
these events, how is that lymphoid cells mediate, actuate, orchestrate their
effects on the wound or its components?
What genes or biochemistry let them exert their adverse, suppressive,
inhibitory, or disruptive dysdynamic effects?
Are the lymphocytes even the chief mischief-makers or just markers
correlated with a more sinister entity?
Are they passive consequences of the problem, dragged secondarily into
the melee, or are they primary culprits?
If they are primary troublemakers, are their effects overt and
audacious, or subtle and surreptitious?
On slide 34 we said “. . . if you are a neighborhood angiocyte, then
that lymphocyte who doesn’t live on your street is probably going to beat you
up . . .” Perhaps that is true, but
perhaps they are just pulling the tires off of your car so you cannot go
anywhere, i.e. not so malicious, but equally effective. That is the key thing about understanding
the dysdynamical physics of this system.
These cell populations with their inhibitory dependencies do not have
to be aggressive and offensive to make trouble. They just have to interfere or disrupt, and
the system can just as likely get disorganized and get onto a go-nowhere
chaotic attractor. It is quite
possible, because one of the quintessential attributes of any chaotic
non-linear system is that subtle little changes can have big dramatic effects. So, in coming up with potential
explanations for what is happening, the following are features that are
observed histologically, with some interpretations and hypotheses. You can see the examples in Part 4, a
gallery of cases, posted on the Arimedica website. Basic anatomical features. Lymphocytes. In the
wound, they look like typical lymphocytes, but in a variety of sizes as they
are reacting to something and accumulating cytoplasm and nucleoplasm to do
their immune functions or become B-cells. Plasma cells. These are
antibody producing B-cells. In the
wound, they look like typical plasma cells.
In the sickest most inflamed or refractory wounds, they can have a
variety of morphological variations and multinucleate forms. Eosinophils. Along with
lymphocytes and plasmacytes, eosinophils are considered part of “chronic
inflammation”. In the wound, they are
typical eosinophils. However,
eosinophils are generally markers of allergy.
Their presence might be an incidental component of chronic
inflammation, but it usually reflects some sort of primary allergic or atopic
state. Drug reactions (due to oral
medications or to wound topicals and dressing materials), atopic and eczematous
dermatoses, and bullous pemphigoid are quite likely the original primary
instigators of the whole situation (just like chronic thrombosis is for those
patients with hypercoagulopathic states and ulcers). Lymphocytes and plasma cells mean immunity,
and eosinophils mean allergy – two similar but different types of host
defense. In the chronic wound,
eosinophils generally represent an primary extrinsic stressor on the wound,
whereas lymphocytes and plasma cells are due to the “intrinsification” of the
process. Lymphoid infiltration.
“Lymphoid” means lymphocytes and plasma cells. In the lymphoid infiltrated wound, both
cells will be found, sometimes almost pure lymphocytes, sometimes almost pure
plasmacytes, and any mix in between.
There is a clear predilection for angioid cells and structures, but
organization gets looser and more admixed as you go higher (earlier) toward
the wound surface. Although incidental
lymphocytes and plasma cells might be seen now and then in young healthy
wounds, they are distinctly NOT a part of normal wound healing, and dense
infiltrates are pathological.
Increasing density of the infiltrates seems to correlate with the
chronicity and refractoriness of each individual wound – worse (harder to
heal) wounds have more. Lymphoid clustering and palisading. It is common to see these cells, especially
plasma cells arranged in straight lines or compact clumps, but this just
represents their alignment along vascular structures. Lymphoid aggregates.
Sometimes lymphoid cells, both lymphocytes and plasmacytes, but especially
lymphocytes will arrange themselves in thick clusters, often appearing
lamellated as they insinuate themselves between layers of angiocytes in a
vessel. These will often persist in
old scars or later in the process, even when things are healing and doing
well. They appear to be chronic
reservoir or standby structures.
Remember that angiocytes NEVER divide preemptively, only in response
to VEGF (or other angiogenic factors) under circumstances of need (growth and
development, injury and wound healing, ischemia, tumors). Lymphoid hyperplasia and activity seems to
be most disarrayed and dynamic in the midst of active wound healing and
inflammation, when angioid cells are themselves their most migratory,
mitotic, and metabolic. As
angiogenesis subsides, mature lymphoid aggregates hug the mature vessels but
seem to have no effect on them. This
all suggests that the lymphocytes can recognize and stand by structures that
they are sensitized to, but that they “see” their specific target and react
only when angiocytes are “awake and walking”, when cytoplasm, nucleoplasm,
and various cell products are exposed. Germinal centers.
Germinal centers (GC) are lymphoid structures where lymphocytes are
becoming hypermutated antigen-specific B-cells, governed by large dendritic
cells in the middle of the GC. I have
not seen true germinal centers, but I have seen some lymphoid aggregates with
large central histiocytic cells which might represent some sort of forme
fruste or transition state. These have
been in the most severely and chronically inflamed wounds. This supports the thesis that longer more
aggressive more sustained acute inflammation is key to the advent of
auto-immune recognition and sensitization. Location of cells.
Looking at the fine structure of the wound, lymphoid cells have a very
strong affinity for angiocytes and vascular structures, everything from
mature non-reactive normal vessels in uninflamed adipose fascias underneath
to individual streaming angiocyte spindles at the top of the wound. They are occasionally aligned with collagen
bundles and fibroblasts. Looking at
the gross architecture of the wound, they seem to be never too far away from
the action. Mature “reservoir”
lymphoid aggregates may be seen in deeper uninflamed areas, but active
infiltrates and clusters seem to be within microns to a few millimeters of
the acute activity. It is common to
see that if there is chronic scar at the base of the wound, with adipose
under that, that the lymphoid infiltration will abate about half way down
through the scar. Deeper infiltrates
seem to have some loose association with primary CVD-CTD diagnoses as opposed
to other primary diagnoses that subsequently become auto-immunized. Lymphocytes in lymphatics.
When infiltrates and aggregates are present, lymphocytes are often
seen packed into lymphatics in the zone of all the action. It is unclear if these are blood borne
lymphocytes that just prefer to then congregate and hang out in the
lymphatics, or if lymphocytes are arriving at the wound via lymphatics. If the are arriving that way, then it
implies that there is some sort of taxis or tropism flagging them down. 3-population mix.
In the uppermost parts of the wound where acute inflammation is most
intense and wound module is least organized, this is where the most intense
and amorphous mixing occurs between the various cell populations. Admixture of equal amounts of acute
inflammation, wound module, and chronic inflammation correlates with the
sickest wounds, and is apt to be seen during active ulceration and divergence
as well as in the most intransigent chaotic wounds. Admixture with necrotic stroma, nuclear and
cytoplasmic and other basophilic debris, and with
monocyte-macrophage-histiocytes is telling evidence of how auto-sensitization
is occurring in the first place. Pathological effects. Destructive effects.
As described above, there is little evidence of direct cell toxicities
or killing by the lymphoid cells. This
is especially obvious in older more mature peri-vascular aggregates away from
acute zones and strata, where lymphoid cells and vessels seem to cooperate or
at least tolerate just fine. It seems
clear that the lymphoid cells have no negative interests or dependencies with
regard to mature inactive angiocytes.
When angiocytes get active, something is revealed to stimulate the
lymphocytes or to attract their attention.
Whatever it is, it is not directly lethal or destructive to the
angiocytes or fibroblasts, and the repair cells and their structures seem to
be architecturally mostly normal. Non-destructive effects.
Non-destructive effects occur in the form of altered architecture and
anatomical disorganization of the wound.
Cell population intermixture is increased, architecture of the newest
vessels can be altered in various ways, (or not), new vascular density can be
altered (too much or too few, usually too few), the plasma protein and
aminoglycan strata can have abnormal thicknesses or ratios. The aminoglycan stratum (which is also the
angioid vertical migration stratum) tends toward the thin side, a crucial
defect, since without a place for angioid cells to develop into mature
vessels, the rest of the process becomes retarded or arrested. Bizarre changes such as excessive angioid
mitoses and presumably accelerated apoptosis are incidental interesting
findings. Matrix and aminoglycan effects. The weakness of the aminoglycan layer in
some of these wounds might or might not have something to do with the
lymphoid aggregates, and if so, then the next question is whether that is a
direct effect of the lymphocytes versus an indirect effect via their effect
on other cells. In the sense that
these parameters are all present together in the impaired wound, they are all
correlated, but what effect lymphoid cells have on the matrix can only be
speculated. I have not yet done alcian
blue stains to look at aminoglycans, but that might be instructive. Effects on angioid cells versus vascular structures. It is easy to think of the immunization
process as making antibodies which will fix complement and invoke killer
T-cells which will then lyse cells and thereby destroy the tissue and create
more ulcer. Perhaps that occurs
sometimes during periods of active ulceration, but our focus here is on the
impaired non-healing chaotic wound that is not actively ulcerating. The lymphoid cells are clearly not directly
lethal to the stromal cells. Perhaps
they would/should be to fungi and bacteria and whatnot, but our own cells are
not getting lysed. It appears much
more likely that lymphoid cells are responding to something that angioid
cells make – some sort of integrin perhaps, some sort of matrix protein or
glycoprotein that binds the vascular locus, some sort of cell surface
receptor, protease, or other exocellular product that angiocytes might
make. The lymphocytes and plasma cells
are certainly getting more stirred up when wound healing is active and
angioid cells are themselves active – transforming, mitosing, migrating,
reassembling – doing all of those angiogenesis things that they normally do
not do in the healthy reserve standby state.
However, to the extent that key products or expressions of angioid
function are affected, this seems to be enough to disorganize overall wound
integration and self-assembly. Effects on stroma materials versus cells. The same rationale applies to possible
effects of the lymphoid cells on other chemicals or materials of the matrix
as opposed to direct toxic effects on angiocytes and fibroblasts. Neutrophil induction.
When a wound is still actively ulcerating and size is enlarging,
something is obviously killing epithelium as well as dissolving mesenchymal
matrix. In biopsies from these
lymphoid wounds where ulceration is still active, neutrophils are seen to be
doing the acute damage. They are
actively infiltrated across basement membrane into the epidermis, dissolving
everything in their path. These
destructive neutrophilic infiltrates are taking place just above
non-destructive lymphoid infiltrates, and it seems that the lymphoid cells
are instrumental in triggering acute inflammation. The hypothesis that the lymphoid chronic
inflammatory cells are having an inhibitory effect on wound module cells
remains correct in the physics-dynamical sense. However, biologically, the direct effect of
the lymphoid cells might be on neutrophils which in turn inhibit the repair
cells. The neutrophils might exert
this influence in two ways. (1) As
discussed in Part 1, acute inflammation has suppressive effects on repair
processes, via a variety of chemical and cellular mechanisms, and neutrophils
stir up acute inflammation. (2)
Neutrophils are making destructive proteases that chew up the matrix. When neutrophil activity is intense, active
ulceration occurs (divergent wound), but at lower levels of activity in the
chaotic non-divergent wound, neutrophils may be continuously degrading the
matrix or inhibiting repair cells just enough to ruin the ability of repair
cells to assemble properly. Non-immunoglobulin effects.
The assumption on seeing plasma cells is that they are immune
competent and making antibodies and thereby killing, defunctionalizing, or
clearing cells. To the extent that
plasma cells are indeed having a direct inhibitory effect on angioid and
fibrous cells, the effect may be by some other unappreciated mechanism. Direct versus indirect effects. To reiterate, from a dynamics point of
view, lymphoid cells are having an inhibitory effect on repair cells. From a biology point of view, that effect
might be direct or it might be indirect through intermediaries or other
chains-of-effect or sub-loops in the system.
The discussions of these last few items simply reinforce how important
is the entanglement of acute inflammation, chronic inflammation, and repair
cells, i.e. the 3-population wound, to inhibit repair by whatever mechanisms
it does so. Dynamical effects. Reservoir functions and standby mode. As discussed above, the lymphoid cells seem
to mature and go into a standby mode around vessels as the tissues mature or
become free from inflammation. These
residual chronic lymphoid aggregates may have no purpose whatsoever, or they
might maintain some state of low level immune or other effects, or they might
be memory cells waiting for some new event to re-activate them. Local versus global effects.
In looking at all of these specimens, from wounds of varying duration,
severity, and refractoriness, from wounds with primary autoimmune disease,
induced wound auto-immunopathy, and a variety of all other primary diagnoses,
I have gotten the sense that lymphoid cells and their aggregates can either be
bred locally and stay local, or they can become global in scope and
effect. (1) The local variety of
infiltrates seems to be in patients whose primary wound diagnosis is
extrinsic, e.g. arterial disease or pressure or radiation. There is no generalized
auto-immunopathy. The effects of
chronicity and cell population admixture to create sensitized lymphocytes is
a process that occurs “right there” and stays right there. The sensitized lymphocytes presumably are
not taking themselves back to the “hive” to be memory cells in some faraway
lymph node or germinal center. The infiltrates are always “within a stone’s
throw” of the wound, representing local breeding of lymphocytes during acute
phases, and those lymphocytes now they stay where they were born, even as the
wound settles down and matures. (2) In
patients who have an a priori CVD-CTD or other autoimmune disorder, and in
patients who secondarily get widespread rheumatoid-lupus like symptoms due to
the wound (e.g. hidradenitis patients), the auto-sensitization of lymphocytes
against stroma, would seem to have a global effect. Whether there are memory cells or not,
specific antibodies or not, whether they remain local or move on to central
lymphoreticular organs are all obvious questions to be asked. This concept might help explain why some of
the intense local lymphoid aggregates now and then have some features which
look like they are trying to grow up and become germinal centers. These might in fact be reservoirs or depots
for local memory cells which can cause persistence of the local wound healing
problems even if the rest of the body is trouble free. This might explain for example why lupus
patients often get local scar problems – cicatritis, panniculitis, or acute
ulceration – when a remote inflammatory event occurs, such as a cold or
pneumonia or urinary infection or trauma.
Even other nominally healthy patients can have similar symptoms, such
as women with silicone breast implants suddenly getting tenderness then
capsular contracture after remote infections or injuries. In those circumstances, remote acute
inflammation seems to be turning on local dormant immune cells which were
bred during the initial injury and wound healing in that particular area. Slow progressive auto-immunization. This is for me one of the most interesting
observations and hypotheses. The most
problematic intrinsically chronic wounds and the patients with the most
active autoimmune diseases seem to have the greatest load of lymphoid cells
in their wounds. However, chronic
wounds of other origins, such as arterial, pressure, radiation, etc. also
have lymphoid infiltrates. Keep in mind
that when you look at benign healthy healing acute wounds, lymphoid infiltrates
are uncommon. However, in our clinic
we see patients with chronic problem wounds that are not healing, and that is
where our biopsies are coming from.
So, it is not surprising that even the simpler healthier wounds for
the more trivial reasons have developed some features of chronicity in the
interval before they started getting good care. At first I thought it peculiar and
unexpected that pressure and arterial wounds were getting lymphoid
infiltrates. However, there are two
other observations about these wounds that are relevant. (1) These otherwise healthy non-autoimmune
patients were also having “clean” wounds with no infiltrates in more acute
areas. (2) The degree or load of the
infiltrates seems to be related to the length or severity of the ulcer. Thus an arterial wound of only a few months
duration might have just a few incidental scattered lymphoid patches, whereas
a pressure ulcer of years duration might have quite a bit more, but still
hardly anything compared to a lupus patient with active ulceration of a
scar. The implication is that stromal
auto-sensitization is occurring locally and that it is a random or stochastic
or statistical function. In any given
time interval, there is a probability S that an element of tissue
within the chronic wound will become sensitized (S is basically the
conversion rate, and it will reflect a variety of local and host factors, so
it will be unique for each patient, wound, and set of circumstances). What is the lymphoid load L in the
wound? First, we normalize the system
by designating wound mass as 1, which means that we are now solving to find
out what fraction of the wound has developed auto-sensitization and lymphoid
infiltrates. There are now two ways to
approach this. (1) We can assume that
the load of lymphoid cells compared to the total mass of the wound is small,
so the available space for sensitization to take place is always roughly
1. In that case, the change in L in a
given interval is just the conversion rate S, so dL/dt = S, which solves
to L = St, a linear zero-order (straight line) solution. i.e. lymphoid
load is directly proportional to time.
(2) Alternatively, if L will be a notable fraction of wound space,
then sensitization can only take place in the remaining areas that are not
yet sensitized. Thus, dL/dT =
S(1-L), which solves to L = 1 – e-St, which is
logarithmic, rising asymptotically to 1.
So, somewhere between a direct
proportionality and a logarithmic curve, there is a time-dependent
uprise in stromal sensitization in any chronic wound. It would seem that in otherwise healthy
patients. this is the kind of sensitization that stays local. It does not imply any type of global
auto-immunopathy, but it does explain why the more chronic (longstanding) a
wound gets, the harder it is for some of them to heal because of the onset of
lymphoid intrinsic chronicity (dynamical). Other items. Biochemical markers of chronicity. Wound research in the past 10 years has
looked at wound chronicity from biochemical and other conventional bioscience
points of view. There are important
differences between acute and chronic wounds.
Matrix proteases and cytokines & growth factors are two general
classes of chemicals which have been studied closely, and genomic profiles
through gene chip analysis have become a leading edge tool in the past 2 or 3
years. These studies all show
substantial differences between healthy and chronic wounds. This would be expected. Acute and chronic wounds are two thoroughly
different states. The findings of increased
proteases and acute inflammatory cytokine profiles in chronic wounds is easy
to understand when you realize that sustained acute inflammation is one of
the necessary dependencies of this system, creating the conditions that lead
to auto-sensitization, and being sustained by both the primary disease or
injury and the intrinsically chaotic state of the wound. Different gene profiles are no surprise due
to sustained disease and injury, sustentation of acute inflammation,
over-activity of the control loop, and the appearance of a third population of cells. Acute versus chronic, converging versus
chaotic, 2-population versus 3-population, Dr. Jekyll versus Mr. Hyde – these
are two very different states, and something has to actuate those
differences, i.e. genes, peptides, other chemicals. It would be very interesting to see
explicitly how much of the variances can be attributed to lymphoid markers,
which should be nearly zero in normal acute healing. The causes of connective tissue disorders. For many years, researchers and clinicians
have mused and hypothesized over the origins of rheumatoid arthritis, lupus,
and the related collagen vascular and connective tissue disorders. Some have ascribed it to an occult pathogen
because it happens in association with prior acute and chronic
infections. Others have ascribed it to
allergic or atopic states. Some to
injury and sustained inflammation. One
of the first “wow, that’s interesting” moments I had as medical student on
clinical rotations was seeing a rheumatoid patient that had a list a mile
long of allergies, to pharmaceuticals and natural allergens. That is common in these illnesses. It is easy to think that their rheumatoid
causes multiple allergies, but most likely it is the other way around, a heightened
state of allergy, atopy, and sustained acute inflammation eventually leads to
stromal auto-sensitization. My first
insights into this issue came from looking at patients, wounds, wound
histology, and the laboratory profiles that we draw on our pathological
wounds. It became clear after a while
that when dealing with hypercoagulable and other micro-occlusive wounds, and also
with “rheumatoid” auto-immune wounds, that sometimes the two classes of
disease were easy to discriminate, but that often they were confusingly
similar to each other, both the patient profiles and the wounds
themselves. Careful history reveals
features of both. Laboratory markers
show unequivocal signs of both categories of disease, as shown in Part
2. When we frequently find Factor V
Leiden in patients with crippling rheumatoid arthritis, that is no
coincidence. It seems that perhaps as
many as 85% of our patients with an apparent primary hypercoagulable state
also have symptoms and laboratory markers of an auto-immune disorder. And as we have developed the thesis here,
the chronic wound is a CVD-CTD in its own right. It was the wound histology that solidified
the appreciation that auto-immune sensitization occurs as a result of the
sustained diffuse micro-thrombosis. What
it all means is that the lupus and rheumatoid class of autoimmune CVD-CTD,
and other auto-immunopathies are all a result of some type of sustained
primary disease and acute inflammation, including thrombosis-hypercoagulability,
infection, allergy-atopy, chronic injury, etc., i.e. the items discussed at
length in Part 2. What is key about
looking at these wounds and their histology is that this thesis can be seen
clearly, item by item, step by step.
This thesis is an integrated explanation of the various reasons why
auto-immunity occurs, accounting for the sciences and observations of
inflammation, immunity, stromal biology, and the clinical and laboratory
profiles of these pathological states and diagnoses. It is necessary now to do large correlative
studies of one class of disease versus another to confirm these
relationships, and to begin looking for the conventional bioscience
laboratory markers of these events. Arimedica.com.
These papers are posted on the Arimedica website, freely available for
appropriate educational, scientific, and clinical use. Part 4 of this series is a companion
presentation with just case studies.
It was not presented as part of the original series due to time
constraints, but the case studies illustrate all of the points in this
epilogue, as well as showing numerous clinical pictures of patients that have
the problems discussed in the first 3 parts. |
|
|
|
|
|
End |
|
|
|
|
|
Abstract (as submitted in advance of the meeting) The Physics and Pathology of
Wounds. Part 3. Chronicity and the Physics of Wound
Failure. Marc E. Gottlieb, MD, FACS Phoenix, AZ The wound module is a transient set of
interacting cells which collectively restore injured tissue to normality, a
fibrous stroma of angiocytes and fibroblasts. Its healthy aggregate behavior
is a well behaved machine, governed by the physics of control systems. A sick
system can result from various extrinsic perturbations, but the core
mechanism of self-sustaining persistent dysfunction, the true intrinsic
disease of wound healing is chronicity itself, the paramount cause being
wound module autoimmunization. This state is disruptive but not fully toxic
or lethal, thus immunopathic wounds have complex behaviors, at times
better-worse-stable-variable, often looking healthy, but always frustrating
as they refuse to cross the finish line. How does one explain such variable
behavior and the differences between normal and chronic-and-pathological
(cap) wounds? Simply stated, intrinsic wound
pathology and chronicity is a dynamical disorder of complex populations. The
physics governing complex behaviors in complex systems is non-linear dynamics
(nld). In addition to control, three aspects of NLD are especially important
to wound pathology. (1) Population logistics. Healthy healing is a sequence
of one-shot self-completing linear events: primary injury & thrombosis
–then– acute inflammation –then– wound module. Pathology creates abnormal
population dependencies (nutrition, starvation, predation, cultivation) and a
new population, chronic inflammation. Non-linear perpetual complexity arises
in the logistics of injury & thrombosis –vs– acute inflammation –vs–
wound module –vs– chronic inflammation –vs– injury & thrombosis. (2)
Cellular automata & self-organization. The “cellular” agents of the wound
module (real biological cells in this case) have a small set of deterministic
rules of interaction with each other.
When allowed to function properly, stromal rebuilding is automatic and
correct. Under pathological
conditions, self-organization, i.e. wound healing is disrupted. (3) Chaos & N-body dynamics. The net
effect is that the wound, a set of several interacting cell populations, has
3 attractors (basins, dynamically stable states or behaviors): convergence
(healing), divergence (ulcerating), and self-sustained chaotic orbits
(chronicity). Basic methods to demonstrate
non-linear dynamics: left, the logistical map of competing populations;
middle, diffusion-limited-aggregation, an example of self-organizing
automata; right, attractors and chaos in the Mandelbrot set of complex-plane
iteration. While seemingly abstract,
these structures are directly correlated with wound events. |
|
|
|
|
|
THE PHYSICS AND
PATHOLOGY OF WOUNDS. PART 3. CHRONICITY AND
THE PHYSICS OF WOUND FAILURE. Original presentation February 22 - 26, 2010, Maui, at the The John A. Boswick, M.D. Burn and Wound Care Symposium 2010 The presentation and related materials are accessible at: arimedica.com Content may be used for non-commercial educational purposes. Content may not be published or used for commercial purposes
without prior license or permission. Contact information is on the slide. Copyright © 2009, Marc E. Gottlieb, MD Revision 01a, February
22, 2010 |
|
|
|
|
|
These presentations and supplementary
materials are all available at www.arimedica.com. The Physics and Pathology of
Wounds. Part 1. The Wound as a System and a Controlled
Machine. Marc E. Gottlieb, MD, FACS Phoenix, AZ The wound is a transient organ of
inter-operating cells, triggered into being by injury and inflammation, then
extinguishing as it completes its repair of injured stroma. It is a system.
Conventional bioscience tends to characterize properties and interactions of
individual or one-versus-another elements within a system, but physics is
required to understand the integrated timewise behavior of whole systems.
Intrinsic wound pathology and chronicity, and wound failure and therapeutics
are easily explained when wounds are seen as a non-linear System (rather than
as a collection of dual-element linear interactions). For normal wound
physiology and for the pathophysiology of altered and failing wounds, the
governing principles are the physics of complex systems: non-linear N-element
dynamics, control science, population logistics, and self-organizing
automata. Understanding wound physics begins by
characterizing normal wound physiology. The wound is a closed-loop
reference-driven non-linear multicontrol system. Sick and altered wounds have
layers of added complexity, but the quintessential intrinsic machinery of
wound healing – the Wound Module of post-inflammatory wound repair –
functions as just a single control loop. When tissues are injured, the Main
Control Loop of physiological wound repair will drive cells to reorganize back
to a repaired stroma. The wound control system is composed
of these elements: The system state is the open wound and its conditions. It
is compared (?) to a reference, normal epithelialized tissue. Variances
generate an error signal in the form of inflammation. This activates
macrophages which are the system controller. They in turn generate a control
signal in the form of cytokines. The controlled load is the group of local
responder cells. Their output are the elements of histogenesis, which modify the
state of the system, which then feeds back to the loop at the summing point.
Any discussion or research of the collective behavior of a wound must
acknowledge this basic control system. |
|
|
|
|
|
These presentations and supplementary
materials are all available at www.arimedica.com. The Physics and Pathology of
Wounds. Part 2. Auto-Immunopathy and the Intrinsic
Disease of Wound Healing. Marc E. Gottlieb, MD, FACS Phoenix, AZ Many chronic wounds result from
disorders extrinsic to the healing process, e.g. pressure or arterial
disease. What then are the intrinsic diseases of wound healing? Compare the
wound to other organs. The quintessence of heart failure is that it is an
inadequate pump, for lung failure it cannot exchange gases. But the wound is
neither pump and pipes, nor bellows and diffusion membrane, nor is it like
any organ with macro-anatomical structure. It is a transient collection of
mutually interacting self-organizing cells.
Stromal angiocytes and fibroblasts (wound cells) have remarkably few
inherent metabolic or genetic faults. Dysfunction of the aggregate population
is almost always the result of deprivation or predation. Adverse states can
be caused by (1) non-targeted exogenous conditions such as arterial ischemia
or repetitive trauma, and (2) targeted damage directed against these cells
and their structures. As will be presented here, predation against the wound
module is due to a state of auto-immunopathy in which lymphoid cells are
sensitized to wound components. Not only does this occur with classic
connective tissue disorders and other well-recognized auto-immunopathies, but
it happens when a wound becomes intrinsically chronic and pathological. Hypercoagulability and other conditions of
persistent thrombosis and acute inflammation are the underlying states that
induce the auto-immunization. Simply put, intrinsic wound pathology and
chronicity is a dynamical disorder of complex populations caused by
auto-immunopathic disruption of the wound module. In these chronic non-healing wound
samples, the vascular locus is infiltrated with immune cells (left, plasma
cells; middle, plasma cells and eosinophils; right, lymphocytes.) On the left, plasma cells are mixed with
the migratory angiocytes (spindles) that are trying to assemble the
wound. At middle and right, chronic
thrombosis due to a primary hypercoagulable disorder is not only present, it
is the root cause of this entire state. |
|
|
|
|
|
These presentations and supplementary
materials are all available at www.arimedica.com. The Physics and Pathology of
Wounds. Part 3. Chronicity and the Physics of Wound
Failure. Marc E. Gottlieb, MD, FACS Phoenix, AZ The wound module is a transient set of
interacting cells which collectively restore injured tissue to normality, a
fibrous stroma of angiocytes and fibroblasts. Its healthy aggregate behavior
is a well behaved machine, governed by the physics of control systems. A sick
system can result from various extrinsic perturbations, but the core
mechanism of self-sustaining persistent dysfunction, the true intrinsic
disease of wound healing is chronicity itself, the paramount cause being
wound module autoimmunization. This state is disruptive but not fully toxic
or lethal, thus immunopathic wounds have complex behaviors, at times better-worse-stable-variable,
often looking healthy, but always frustrating as they refuse to cross the
finish line. How does one explain such variable behavior and the differences
between normal and chronic-and-pathological (cap) wounds? Simply stated, intrinsic wound
pathology and chronicity is a dynamical disorder of complex populations. The
physics governing complex behaviors in complex systems is non-linear dynamics
(nld). In addition to control, three aspects of NLD are especially important
to wound pathology. (1) Population logistics. Healthy healing is a sequence
of one-shot self-completing linear events: primary injury & thrombosis
–then– acute inflammation –then– wound module. Pathology creates abnormal
population dependencies (nutrition, starvation, predation, cultivation) and a
new population, chronic inflammation. Non-linear perpetual complexity arises
in the logistics of injury & thrombosis –vs– acute inflammation –vs–
wound module –vs– chronic inflammation –vs– injury & thrombosis. (2)
Cellular automata & self-organization. The “cellular” agents of the wound
module (real biological cells in this case) have a small set of deterministic
rules of interaction with each other.
When allowed to function properly, stromal rebuilding is automatic and
correct. Under pathological
conditions, self-organization, i.e. wound healing is disrupted. (3) Chaos & N-body dynamics. The net
effect is that the wound, a set of several interacting cell populations, has
3 attractors (basins, dynamically stable states or behaviors): convergence
(healing), divergence (ulcerating), and self-sustained chaotic orbits
(chronicity). Basic methods to demonstrate
non-linear dynamics: left, the logistical map of competing populations;
middle, diffusion-limited-aggregation, an example of self-organizing
automata; right, attractors and chaos in the Mandelbrot set of complex-plane
iteration. While seemingly abstract,
these structures are directly correlated with wound events. |
|
|
|
|
|
|
|
|
|
|